IJCRR - 3(4), April, 2011
Pages: 83-94
Print Article
Download XML Download PDF
GENETIC MECHANISMS OF DROUGHT TOLERANCE IN MAIZE HYBRIDS DEVELOPED FROM SCREENING OF 100 MAIZE LINES
Author: Roshni Vijayan, A.Kalamani, G. Nallathambi
Category: General Sciences
Abstract:Drought is a major constraint in the cultivation of crops especially in tropical areas. Drought tolerance studies in maize help to understand the parameters which are associated with drought stress in the crop. This research was conducted to analyze the impact of drought on yield of maize (Zea mays L.). The development of drought tolerant hybrids for rainfed and moisture stress conditions with emphasis on using genotypic and phenotypic data was another objective. The initial study started with 100 genotypes from which the 10 best genotypes (lines) were selected for drought tolerance studies. Screening was carried out using physiological and phenotypic data.
Thirty hybrids were developed from the 10 lines and 3 testers (locally adapted varieties) utilizing a LINE X TESTER analysis. Parents and hybrids were phenotypically assessed in two field conditions: irrigated and moisture stress. Results showed that hybrid IBET IE 1253-8 X UMI 61 was best under normal irrigation and IBET IE 1256-6 X COH (M)5 was best under moisture stress. Taking both fields together, the best hybrid was IBET IE 1253 X UMI 61 which averaged 6.4t/ha. The best parental lines for both conditions were COH (M) 5 and Hy R'06 6143-16. Results support the fact that yields are low when maize is subjected to drought stress. The best
hybrid was equal to the local variety under both irrigated and moisture stress condition. Drought tolerant maize hybrids can help to improve productivity in drought stressed areas.
Keywords: Drought, line X tester, moisture stress, gca, sca
Full Text:
INTRODUCTION
Maize (Zea mays.L.) is the most important crop next to wheat and rice in the world agricultural economy. It is important for food, animal feed and industrial utilization. It is the crop of the future as mentioned by Dr. Norman E. Borlaug (Dahiya 2008). Maize can play a vital role in ensuring food security as well as nutritional security for developing countries and the world as a whole. According to the Food and Agriculture Organisation (2003) report, out of 593 million tons of maize produced on 142.3 million hectares globally, 17 per cent is used as human food and 66 per cent as animal feed. Drought is a major constraint to maize production in all areas where there is no adequate rains or irrigation. Global climate change is considered to be underway and is expected to result in a long-term trend towards higher temperatures, greater evapo-transpiration and an increased incidence of drought in many regions of the world (Hillel and Rosenzweig. 2002). Water shortage in India by 2020 is expected to be severe, as the per capita availability of water per person becomes less than 1,000 cubic meters. Water shortage is important, as 85 per cent of water is used for agriculture, 10 per cent for industry and 5 per cent for domestic use (outlookindia. 2009). Above mentioned trends emphasis the fact that competition for water will increase in future with domestic and industrial demands meaning less water for agriculture and irrigation. The expansions of cultivation of maize into marginal production areas are generating increasingly drought-prone maize production environments. The grain loss in maize due to drought in the tropics averages 24 million mega grams per year (Wilkinson 1992). Climate change will further increase the chances of drought. Maize hybrid breeding objectives depend on various factors like the requirements of farmers, market forces, production levels, constraints and crop ecology. However, the most important breeding objective is increased grain yield. The average annual yield loss in maize due to drought is estimated to be 17% in the tropics (Wilkinson 1992). Drought tolerance is not a simple character governed by one or two genes but controlled by a number of morphophysiological characters independently controlled by more than two genes (Fukai and Cooper. 1995). The major problem in drought tolerance breeding is the poor understanding of genetics and the inheritance of drought tolerant traits, and lack of understanding of the relationship between the physiological traits in drought tolerance and plant productivity under stress (Wilkinson 1992). Improvement of drought resistance in high yielding genotypes can be achieved by the incorporation of morphological and physiological mechanisms of drought resistance in new lines through breeding programs. The use of genetics to improve drought tolerance and to provide yield stability is an important part of the solution to stabilizing global maize production. Breeding genotypes suitable for both irrigated and drought conditions will be useful to farmers and to industries. This research uses physiological and morphological drought tolerant mechanisms for the development of drought tolerant hybrids. The objective of the study was to provide useful information for breeders in developing drought tolerant maize hybrids. The activities of the research comprised of three parts: i) Screening of the germplasm for drought tolerance ii) Line x Tester analysis.
MATERIALS AND METHODS
The present investigation was carried out from 2007- 2009 at the Department of Millets, Centre for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore, India (11 North latitude, 77 East longitude). Soils for the experiment were black cotton soils (vertisols) with a loamy texture for the irrigated treatment and a clay loam for the induced moisture stress treatment. During the experiment the mean annual rainfall was 17.7mm received in 4 rainy days. The mean maximum and minimum temperatures were 30.2 C and 21.5 C, respectively.
Screening of maize genotypes for drought tolerance
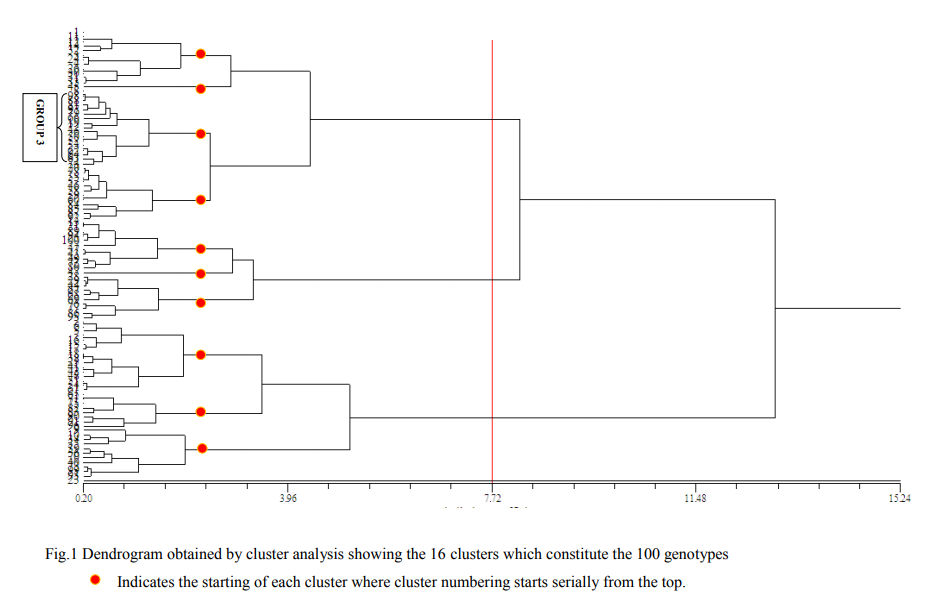
A hundred lines obtained from the maize germplasm collection at the Department of Millets, Centre for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore were screened for drought and identification of tolerance traits. Initial Screening was done in a greenhouse using relative water content, chlorophyll content values and specific leaf area data. Seeds were sown in 250 ml paper cups filled with garden soil, peat and sand in 1:1:3 ratios. A completely randomized design (CRD) with 3 replicates, with 10 plants per treatment was used. All 3 measurements were taken from five plants randomly selected from each replication on the 30th day after seedling emergence. The Chlorophyll meter from Minolta (SPAD model 502 of Minolta, Japan) was used to measure chlorophyll values from the newly emerged leaf on either side of the midrib at a point three - fourths of the way from the base to leaf tip. Specific leaf area is a measure of the leaf area of the plant to leaf dry weight. Matured leaves after 20 days of emergence were collected and leaf area was measured using a LI-3100C Area Meter and leaf dry weight was taken after oven drying for 8 hours at 80C The relative water content was estimated by taking 25 leaf discs of 1 cm diameter from 25 day old leaves using a punch machine and fresh weight of the leaf discs was taken. The leaf discs were then immersed in water at 25C. After 4 hours the turgid weight of the discs was measured. The leaf discs were oven-dried for 8 hours at 80C and the dry weight taken. The formula proposed by (Barrs and Weatherley 1962) was utilized to derive RWC values. For the 100 genotypes, the means of the 3 measured traits were used to do cluster analysis by numerical taxonomy techniques, using NTSYS-pc package, version 2.01, (Rohlf. 1997). An Unweighted Pair-Group Method of the Arithmatic (UPGMA) (Sokal and Michener. 1958) average clustering procedure was employed to construct the dendrogram. Based on the groupings in the dendrogram, 34 genotypes were selected for further studies.
Line x Tester analysis
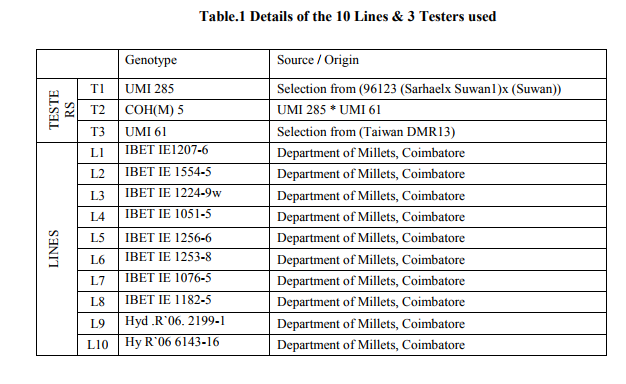
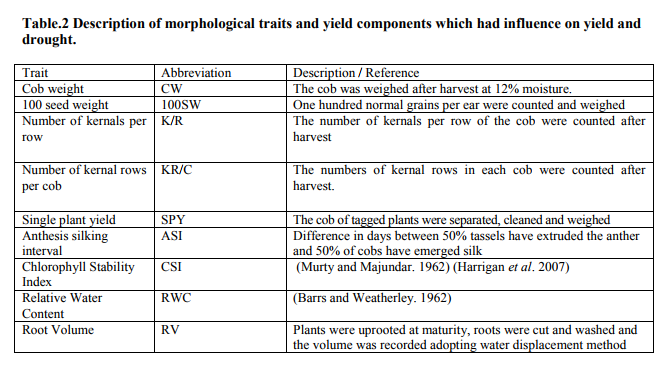
The selected 34 genotypes were taken to a field crossing block during June 2008. Each genotype was raised on a ridge spaced 60 cm apart, five meter long, and 30 cm between plants; sowing of genotypes was at three dates, 10 days apart, for synchronization of flowering. From 34, 10 were selected for the line x tester analysis based on the anthesis to silking interval of 1-2 days. The ten lines selected were crossed with three testers (locally adapted varieties) individually in a line x tester model (Kempthorne. 1957) to obtain 30 hybrids. All thirteen parent lines (10 lines and 3 testers) (Table1) along with the 30 hybrids were evaluated from December 2008 to February 2009, under irrigated and induced moisture stress conditions in two different fields. The moisture stress was created artificially by withdrawing irrigation a week before flowering started for 28 days and another field grew the same lines under normal irrigated conditions with 10 irrigations spaced 10 days apart. Each entry was grown in two rows of five-meter long using a randomized block design(RBD) replicated thrice; spacing was 60 x 30 cm. Recommended agronomic and plant protection practices were followed to maintain healthy growth. Observations were recorded on five plants selected randomly from each entry and replication. The parents and F1 hybrids were examined for morphological traits and yield components whose genes were found to have an impact on drought stressed output as grains. The yield components which were responsible for yield were cob weight, 100 seed weight, number of kernels per row, number of rows per cob, and single plant yield. The morphological traits which showed an influence on drought tolerance of the crop includes, anthesis silking interval (ASI), root volume, chlorophyll stability index (CSI), and relative water content (Table 2).
Statistical Analysis
The data was subjected to statistical analysis using the INDOSTAT statistical package developed by Indostat Services, Hyderabad, India. Analysis of variance for all characters was determined (Panse and Sukhatme. 1961). The mean values were used to estimate heterosis per cent under three categories of mid parent, better parent and standard parent (Fonesca and Paterson. 1968).
RESULTS
Screening of maize genotypes for drought tolerance
Means of 3 measured traits, chlorophyll meter reading, relative water content and leaf area were used to do cluster analysis and develop a dendrogram (Fig 1). This allowed the hundred genotypes selected for drought testing to be grouped into eleven major clusters (Fig.1) and 34 genotypes were selected, with 3 each from 8 groups, 1 each from group 2 and 6 and rest 8 from the group 3 which was the largest group. The 34 genotypes were selected as drought tolerant genotypes for further study which are diverse from each other. These genotypes were planted in field. The 10 genotypes (lines) which had an ASI of 1-2 days were further selected for Line X Tester analysis, since ASI is the major criteria for selection of lines with drought tolerance in maize.
Line X Tester Analysis
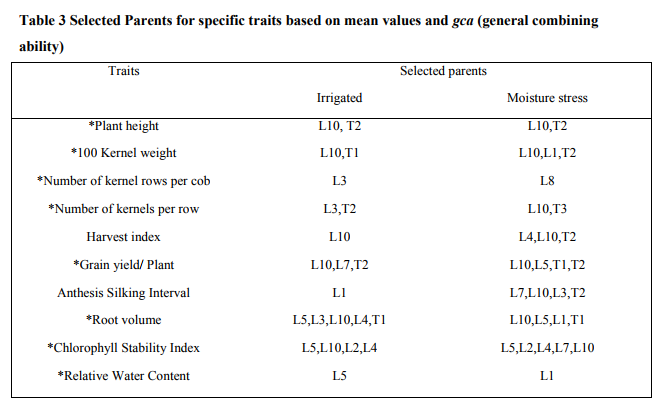
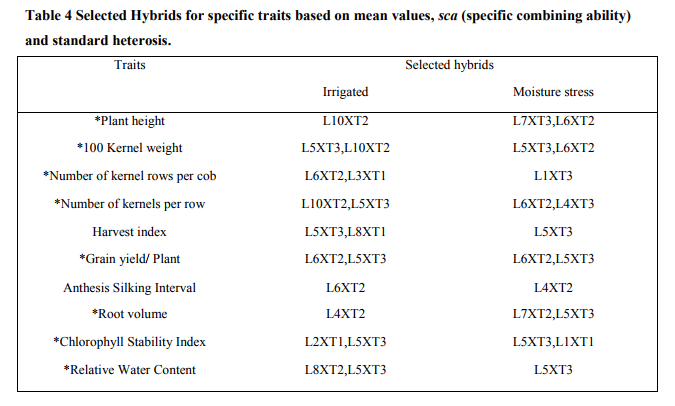
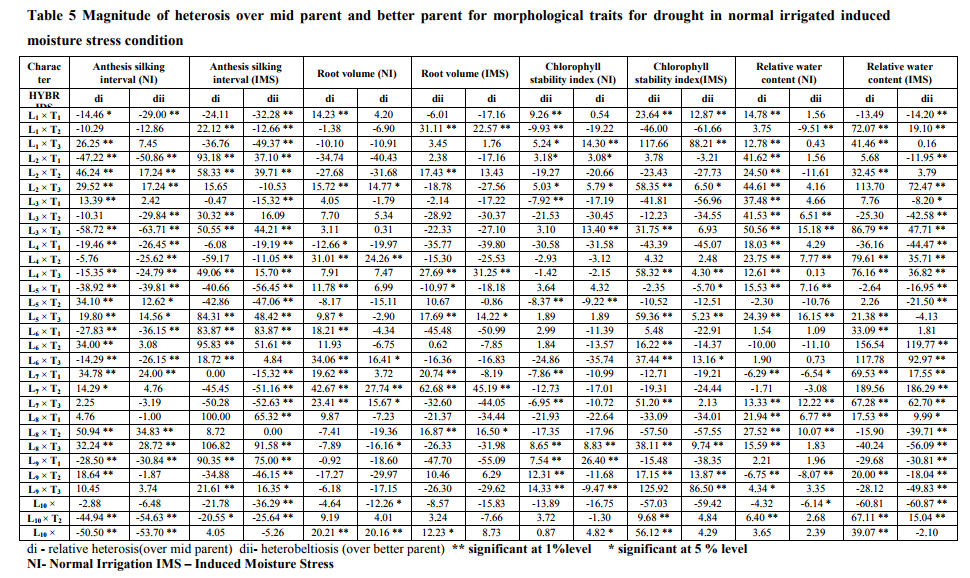
The present investigation tested ten inbred lines as female parents and three inbreds as male parents for their genetic potential in breeding program for grain yield under irrigated and stress conditions. The parents which possess desirable genes were identified by comparing the mean values and general combining ability (gca). The line L10 and tester T2 was found to perform best (Table3). The magnitude of heterosis was studied for characters that affected yield and drought tolerance (Table 5). Since yield is a complex trait, knowledge of the association of different components with yield is necessary. For the expression of maximum yield, all genes which contribute to yield should have a positive heterosis for the exploitation of hybrid vigor in the development of hybrids (Table 4). The major traits which have a genetic contribution to yield showing significant effects are cob weight, 100 kernel weight, number of kernels per row, number of kernel rows per cob, single plant yield. All these traits were showing positive heterosis in the superior F1 hybrids developed. Not much variation was noticed in normal irrigated or induced moisture stress developed hybrids in terms of hybrid vigor. This has lead to the selection of a particular hybrid which performs well in both conditions. A positive heterosis in chlorophyll stability index, relative water content and negative heterosis in traits of root volume and anthesis silking interval is desired to exploit heterosis in development of drought tolerant hybrids (Table 4).This was noticed in the case of both the conditions of normal irrigation as well as induced moisture stress. There was maximum contribution of crosses resulted from lines x testers to total variance for grain yield per plant, 100 seed weight and cob weight, CSI, root volume followed by contribution of female parents for ASI, and plant height. In normal irrigated condition, the hybrid L5XT3 was best suited to heterosis breeding for cob weight, 100 kernel weight, number of kernels per row, and single plant yield. The hybrid L10XT2 was suited for number of kernels per row and 100 kernel weight. For drought prone environments, the hybrids L6XT2 was suited to heterosis breeding for 100 kernel weight, number of kernels per row and single plant yield. The hybrid L5XT3 was suited for root volume. The hybrid L1XT3 was best for number of kernel rows. In induced moisture stress situation, the hybrid L6 X T2 (IBET IE 1256-6 X COHM5) was performing better with positive significant value for grain yield and number of kernel rows per cob while in the irrigated condition, the hybrid L5 X T3 (IBET IE 1253-8 X UMI 61) recorded positive significant values for grain yield and drought related traits of root volume, chlorophyll stability index and relative water content and was the best hybrid for irrigated condition. In both induced moisture stress and normal irrigation, the hybrid L5 X T3 (IBET IE 1253-8 X UMI 61) recorded positive significant values for grain yield per plant and was the best hybrid for both conditions. While considering the various traits in both conditions of induced stress and normal irrigation, the hybrid L5 X T3 expresses a positive significant value for grain yield per plant, which can happen only when the genes which contribute to maximum yield are expressed well in the hybrid along with the desirable environmental effects. The hybrid L5 X T3 (IBET IE 1253-8 X UMI 61) did not show a wide difference in grain yield of 6.4t/ha, 6t/ha for irrigated and induced stress conditions respectively. In the L5 X T3 hybrid both the parents has positively significant gca effects for most of the traits.
DISCUSSION
Screening of the germplasm for drought tolerance From the dendrogram data, it is clear that physiological parameters play a vital role in the study of drought tolerance. The selection of suitable plants from a germplasm collection using specific physiological traits is a viable way for crop improvement for water stress tolerance (Kiani et al. 2007, Reynolds et al. 2005, Tambussi et al. 2007) The initial selections of 34 inbred lines from the 16 clusters in the dendrogram (Fig 1) were based on the physiological traits related to drought. Hence a simple method of screening maize lines using three potential physiological selection criteria, relative water content, chlorophyll content and specific leaf area were tested in the present study. The clusters indicate the diverse nature of the inbred lines used for development of drought tolerant hybrids. (Burke. 2001, Srikanthbabu et al. 2002) expressed that genetic variability for water stress response can only be expressed when plants are exposed to water stress. The higher water stress tolerance is due to expression of water stress-responsive genes that can be translated into certain physiological phenomena such as maintenance of relative water content and chlorophyll content (Bruce et al. 2002, Waseem et al. 2006). Selection of inbreds with the physiological parameters in the initial stage of seedlings itself has an impact on the final development of hybrids since physiological traits are found to have its impact in drought tolerance.
Line x Tester analysis
The ten inbred lines identified as female parents and three testers as male parents, according to the Line X Tester analysis, the genetic potential and the nature of the gene action of the genes involved was assessed. The best parents L10 (Hy R`06 6143-16) and T2 (COH (M) 5) identified had a better general combining ability, but those were not able to produce the better hybrids. The reason could be the good combiners will not result in specific combiners leading to better hybrids. The F1 hybrids which were evaluated for grain yield under irrigated as well as in stress conditions (Ganunga. 2007). In the present study, the magnitude of heterosis, and general and specific combining abilities were studied for drought specific characters (Dadheech and Joshi. 2007 ; Fonesca and Paterson. 1968). Since yield is a complex trait, knowledge of the association of different components with yield like cob weight, number of kernel rows per cob, number of kernels per cob and interrelation among themselves is useful. Per cent heterosis over mid parent and better parents were estimated to know the possible gene action and to exploit heterosis for drought associated traits and yield. Parents with high mean values like L10 and T2 are preferred for using in hybridization program as they are expected to produce desirable segregants (Frova et al. 1999). Evaluation of hybrids in F2 generation becomes necessary to consider whether a hybrid can be used on a commercial basis or could be utilized in a breeding program. The hybrids can be evaluated based on their mean value and degree of heterosis. For cob weight, 100 kernel weight, number of kernels per row, and single plant yield the hybrid L5XT3 recorded the highest value under normal irrigated condition. This shows that genes contributing these traits were able to express better in this hybrid. The specific combining ability of this hybrid is better. While comparing the induced moisture stressed field, the variation observed is due to the impact of stress condition. In a stressed situation certain genes are expected to express well resulting in overdominance of the genes related to drought resulting in another hybrid to express better. In induced moisture stress the hybrid which performed best was L6 X T2. While comparing both hybrids L6XT2 AND L5XT3, the parents involved were different in both. For obtaining good specific combining crosses, good combiners should come together, but this is not a rule for all crosses. Sometimes two poor combiners may yield good specific combination due to epistatic gene action. The crosses with poor combiners will yield transgressive segregants in segregating generations.
CONCLUSION
The development of drought tolerant maize hybrids can help to improve productivity in drought stressed areas. From the research the F1 hybrids developed had the presence of drought tolerant genes. Now when we need to extend the research for F2 generation where we will be able to see the segregating pattern of the genes and also will be able to select the best from the F2 for further development of hybrids for commercial cultivation.
ACKNOWLEDGMENTS
I thank Professor Peter Hobbs, Cornell University, Ithaca, New York, USA for his valuable suggestions in the development of this manuscript.
References:
1. Barrs, H.D. and Weatherley, P.E. (1962) A reexamination of the relative turgidity technique for estimation of water deficits in leaves. Australian Journal of Biological Sciences. 15: 413-428.
2. Bruce, W.B., Edmeades, G.O. and Barker, T.C. (2002) Molecular and physiological approaches to maize improvement for drought tolerance. J Exp Bot. 53: 13-25.
3. Burke, J.J. (2001) Identification of genetic diversity and mutations in higher plant acquired thermotolerance. Physiol Plantarum. 112: 167-170.
4. Dadheech, A. and Joshi, V.N. (2007) Heterosis and combining ability for quality and yield in early maturing single cross hybrids of maize (Zea mays L.). Indian J Agric Res. 41: 210- 214.
5. Dahiya, S.B. Coarse cereals need government support. TribuneIndia...AgricultureTribune. 2009.
6. Fonesca, S. and Paterson, F.L. (1968) Hybrid vigour in seven parental diallel cross in common wheat. (Triticum aestivum L.). Crop Science: 85-88.
7. Frova, C., Krajewski, P., Di Fonzo, N., Villa, M. and Sari-Gorla, M. (1999) Genetic analysis of drought tolerance in maize by molecular markers. I. Yield components. Theor Appl Genet. 99: 280-288.
8. Fukai, S. and Cooper, M. (1995) Development of drought-resistant cultivars using physiomorphological traits in rice. Field Crops Res. 40: 67-86.
9. Ganunga, R.P. (2007) Genotypic and phenotypic chacterization of maize test cross hybrids under stressed and non stressed conditions.
10. Harrigan, G.G., Stork, L.G., Riordan, S.G., Ridley, W.P., Macisaac, S., Halls, S.C .et al. (2007) Metabolite analyses of grain from maize hybrids grown in the United States under drought and watered conditions during the 2002 field season. J Agric Food Chem. 55: 6169-76.
11. Hillel, D. and Rosenzweig, C. (2002) Desertification And Its Relation To Climate Variability And Change. Adv Agron. 77: 2- 39.
12. Kempthorne, O. (1957) An introduction to genetic statistics. Wiely, New York.
13. Kiani,P. S , Grieu, P , Maury, P. , Hewezi, T. , Gentzbittel, L. and Sarrafi, A. (2007) Genetic variability for physiological traits under drought conditions and differential expression of water stress-associated genes in sunflower (Helianthus annuus L.). TAG.Theoretical and applied genetics.Theoretische und angewandte Genetik. 114: 193-207.
14. Murthy,K.S. and Majundar,S.K. (1962) Modifications of the techniques for determination of chlorophyll stability index in relation to studies of drought resistance in rice. Curr. Sci., 31:470-471.
15. outlookindia. (2009) news.outlookindia.com | 'India to Face Severe Water Stress by 2020'.
16. Panse, V.G. and Sukhatme, P.V. (1961) Statistical Methods for Agricultural Workers.
17. Reynolds, M.P., Mujeeb-kazi, A. and Sawkins, M. (2005) Prospects for utilising plant-adaptive mechanisms to improve wheat and other crops in drought- and salinity-prone environments. Ann Appl Biol. 146: 239-259.
18. Ribaut, J.M and Ragot, M. (2007) Markerassisted selection to improve drought adaptation in maize: the backcross approach, perspectives, limitations, and alternatives. J Exp Bot. 58: 351-60.
19. Rohlf, F.J. (1997) NTSYS-PC, E. Setauket, NY. . 2.1 ed, Exeter Software.
20. Sokal, R.R. and Michener, C.D. (1958) A statistical method for evaluating systematic relationships. . University of Kansas, Lawrence.
21. Srikanthbabu, V., Ganeshkumar, Krishnaprasad, B.T., Gopalakrishna, R., Savitha, M. and Udayakumar, M. (2002) Identification of pea genotypes with enhanced thermotolerance using temperature induction response technique (TIR). J Plant Physiol. 159: 535-546.
22. Tambussi, E.A., Bort, J., Guiamet, J.J., Nogues, S. and Araus, J.L. (2007) The Photosynthetic Role of Ears in C3 Cereals: Metabolism, Water Use Efficiency and Contribution to Grain Yield. Crit Rev Plant Sci.
26: 1-16. 23. Tuberosa, R., Salvi, S., Giuliani, S., Sanguineti, M.C., Bellotti, M., Conti, S. et al. (2007) Genome-wide Approaches to Investigate and Improve Maize Response to Drought. Crop science. 47: S120.
24. Waseem, M., Athar, H. and Ashraf, M. (2006) Effect of salicylic acid applied through rooting medium on drought tolerance of wheat. Pakistan journal of botany. 38: 1127- 1136.
25. Wilkinson, D. (1992) Proceedings of the Forty-Seventh Annual Corn and Sorghum Research Conference 1992: [held in Chicago, Illinois, December 9 - 10, 1992]. . American Seed Trade Assoc., Washington, DC.
|
 IJCRR
IJCRR 





 This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License





