IJCRR - 6(21), November, 2014
Pages: 26-30
Date of Publication: 11-Nov-2014
Print Article
Download XML Download PDF
LEPTIN - IT'S ROLE IN ENERGY HOMEOSTASIS
Author: Dipti Mohapatra, Prakash K. Sasmal, Nibedita Priyadarsini, Ellora Devi, Priyambada Panda
Category: Healthcare
Abstract:Leptin (from the Greek leptos, meaning thin) is a 167-amino acid protein hormone with important effects in regulating body weight, metabolism and reproductive function. Leptin, an adipose tissue derived hormone relays information about the peripheral energy reserve and their availability to the brain. This review article is on understanding the physiology of leptin and its role in energy homeostasis thereby controlling obesity, neuroendocrine function, energy metabolism and insights into emerging clinical applications. It also addresses the possible therapeutic uses of leptin in humans.
Keywords: Leptin receptor, Obesity and receptor resistance
Full Text:
INTRODUCTION
Obesity is a rising menace globally, especially in the rapidly developing countries. The weight of the individual is maintained by a balance between the energy intake and expenditure. It is because of the easy availability of high calories diet and lack of physical activity due to sedentary lifestyle that obesity is an emerging problem. The excess energy is stored in the form of triglycerides in the adipose tissue. An interesting neuroendocrine system controls the energy balance in the body by constantly monitoring energy storage, availability, and consumption. Adipose tissue and the brain are the two main components of this neuroendocrine system. The brain controls energy homeostasis and body weight by integrating various metabolic signals released in peripheral tissues. One of the key components is leptin, an adipose tissue derived hormone that conveys critical information about peripheral energy storage and availability to the brain. The term leptin is derived from the Greek word leptos, meaning “thin”. Leptin is a protein approximately of 16 kDa in mass and encoded by the obese (ob) gene1 . In human beings the gene for leptin is located on chromosome 7. Leptin, the genetic defect of ob/ob mice was first described in the 1950s as the spontaneous mutation that causes a severe obese phenotype due to both overeating and decreased energy expenditure. The gene was named ob and the obese mice carrying the mutation were called ob/ob mice2 . Leptin is a 167-amino acid protein hormone with important effects in regulating body weight; metabolism and reproductive function3 . Leptin is expressed mainly by adipocytes, which fits with the idea that body weight is sensed as the total mass of fat in the body4 . Leptin in smaller amounts of are also secreted by cells in the epithelium of the fundus of the stomach, skeletal muscle, liver, placenta, heart, in granulose and cumulus oophorus cells of the human ovaries, human mammary gland and gastric epithelium5-7. The Adipocyte Brain axis: The adipose tissue–derived hormone leptin is produced in proportion to the body fat stores. Circulating leptin serves to communicate central nervous system (CNS) the state of body energy repletion in order to suppress food intake and permit energy expenditure 8-10. Leptin decreases appetite and increases energy expenditure through sympathetic activation, which consequently decreases adipose tissue mass and body weight through its action on the hypothalamic nuclei (Fig. 1). To regulate energy balance in humans the hormone levels are decreased during fasting and increased after several days of overfeeding. So leptin is considered a homeostatic hormone regulating food intake and body weight. Due to latter homeostatic control mechanism, leptin is an anti-obesity hormone, based on the hypothetical fact that high leptin levels would prevent the occurrence of obesity. Unfortunately, this is not the case, and so the strong correlation between serum leptin levels and body fat mass found in obese individuals now suggests the existence of an endogenous leptin-resistant mechanism in obesity 11. The Leptin Resistance as the important cause for obesity: Leptin was also proposed to be a satiety factor because a defect in leptin molecule or receptors led to overeating and obesity. Circulating leptin levels are directly proportional to adipose tissue mass12. High serum levels of leptin signal the presence of sufficient energy stores to sites in the central nervous system, which respond by reducing appetite and increasing energy expenditure, preventing severe obesity. Therefore, leptin signals the nutritional status from the periphery to the area of the brain involved in the homeostasis of energy balance. However, the primary function of leptin may not be as a satiety factor. Leptin treatment at physiological levels reduces eating (and increases energy expenditure) by ob/ob mice to the levels of normal mice, but it does not cause satiety (end of eating). Higher doses of leptin are required to decrease food intake in normal animals13. The same relationships are true in humans with the ob gene defect and normal humans. Leptin levels are also modulated acutely. For example, leptin levels change rapidly with feeding or fasting disproportionately to the changes in fat depot. Therefore, leptin is not just a read out of the fat stores14. Leptin is expressed predominantly by adipocytes, which fits with the idea that body weight is sensed as the total mass of fat in the body, it is a key mediator in the regulation of food intake and energy expenditure. Whereas women have higher leptin concentrations, even after correction for body fat mass, in both genders the subcutaneous fat depot seems to be a stronger predictor of leptin levels than intra-abdominal fat15.Body adiposity has been shown to be a major determinant of circulating leptin. Leptin appears to function more as a signal of recent energy balance than as an “adipostat.”16. Leptin’s effects on body weight are mediated through hypothalamic centers that control feeding behavior and hunger, body temperature and energy expenditure17. Decreased hunger and food consumption is mediated at least in part by inhibition of neuropeptide Y synthesis. Neuropeptide Y is a very potent stimulator of feeding behavior. Increased energy expenditure is measured as increased oxygen consumption, higher body temperature and loss of adipose tissue mass. As expected, injections of leptin into db/db mice, which lack the leptin receptor, had no effect. When leptin was given to normal mice, they lost weight, showed profound depletion of adipose tissue and manifest increases in lean mass. The mechanisms by which leptin exerts its effects on metabolism are largely unknown and are likely quite complex. In contrast to dieting, which results in loss of both fat and lean mass, treatment with leptin promotes lipolysis in adipose tissue, but has no apparent effect on lean tissue18. Leptin has also been implicated in roles as diverse as the regulation of the immune system and respiratory function. In addition, ongoing studies are clarifying the central nervous system (CNS) mechanisms by which leptin exerts its effects on energy balance as well as suggesting important actions for leptin in regulating peripheral metabolic fluxes19-23. Adequate leptin levels also permit energy expenditure in the process of reproduction and growth and similarly regulate the autonomic nervous system, other elements of the endocrine system and the immune system 24-25.
LRs and sites of leptin action
The effect of leptin is mediated by receptors (Ob-R). The leptin receptor contains an extracellular ligand-binding domain, a single transmembrane domain and a cytoplasmic signaling domain and is member of the interleukin (IL)-6 receptor families of class 1 cytokine receptors 26. Leptin receptors are highly expressed in areas of the hypothalamus known to be important in regulating body weight, as well as in T lymphocytes and vascular endothelial cells (Fig. 2). There are multiple LR isoforms, all of which are products of a single lepr gene27-28. The lepr gene contains 17 common exons and several alternatively spliced 3¢-exons. In mice, the six distinct LR isoforms that have been identified are designated LRa–LRf. In all species, LR isoforms can be divided into three classes: secreted, short and long. The secreted forms are either products of alternatively spliced messenger RNA (mRNA) species (e.g. murine LRe, which contains only the first 14 exons of lepr) or proteolytic cleaveage products of membrane bound forms of LR. These secreted forms contain only extracellular domains that bind circulating leptin, perhaps regulating the concentration of free leptin. Short form LRs (LRa, LRc, LRd and LRf in mice) and the long form LR (LRb in mice) contain exons 1–17 of lepr and therefore have identical extracellular and transmembrane domains as well as the same first 29 intracellular amino acids, but diverge in sequence thereafter due to the alternative splicing of 3¢ exons. Short form LRs contain exons 1–17 and truncate 3–11 amino acids after the splice junction. LRc-, LRd- and LRf specific sequences are not well conserved among species. However, LRa (the most abun-dantly expressed isoform) is reasonably well conserved, as is LRb, which has an intracellular domain of approximately 300 residues. LRb is crucial for leptin action. Indeed, the originally described db/db mice lack LRb (but not other LR forms) as a consequence of a mutation that causes mis-splicing of the LRb mRNA; these mice display a phenotype that is indistinguishable from that of db3J/ db3J mice (which are deficient in all LR isoforms) and of leptin-deficient ob/ob animals 29. The function of shortform LRs is less clear, although proposed roles include the transport of leptin across the blood-brain barrier and the production of circulating LR extracellular domain to complex with leptin. Many of the effects of leptin are attributed to effects in the CNS, particularly in the basomedial hypothalamus, a site of high LRb mRNA expression 30-33. Here, leptin acts on neurons that regulate levels of circulating hormones (e.g. thyroid hormone, sex steroids and growth hormone) 34. Leptin action on these hypothalamic neurons also regulates the activity of the autonomic nervous system, although direct effects of leptin on LRb-containing neurons in the brainstem and elsewhere probably also have an important role 35. The effects of leptin on the immune system appear to result from direct action on T cells that contain LRb 36. Leptin might also regulate glucose homeostasis independently of effects on adiposity; leptin regulates glycemia at least partly via the CNS, but it might also directly regulate pancreatic b cells and insulin-sensitive tissues 37-40. Like other cytokine receptors, LRb does not contain intrinsic enzymatic activity but instead signals via a noncovalently associated tyrosine kinase of the Jak kinase family (Jak2 in the case of LRb).
A. Jak-STAT pathway
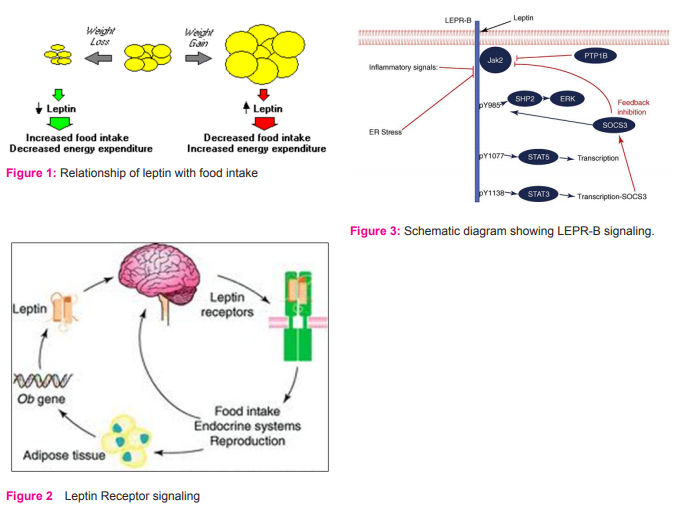
LEPR-B appears to be competent for intracellular signal transduction and is critical for energy homeostasis. LEPR-B upon leptin binding to its extracellular domain, undergoes a conformational change to activate its associated Jak2 tyrosine kinase 41. Activated Jak2 promotes the tyrosine phosphorylation of several intracellular residues on LEPR-B (also on Jak2 itself) and each tyrosine phosphorylation site recruits a specific set of downstream molecules to promote specific intracellular signals. LEPRB contains three distinct tyrosine phosphorylation sites: Tyr985, Tyr1077 and Tyr1138 42. Tyr1138 recruits signal transducer and activator of transcription (STAT 3), a latent transcription factor, which subsequently becomes tyrosine phosphorylated (pSTAT3) by Jak2, enabling its nuclear translocation and promoting its transcriptional effects. Detection of pSTAT3 is used as an important bioassay of LEPR-B signaling in vivo 43. Similarly, Tyr1077 recruits and mediates the phosphorylation and activation of a related transcription factor, STAT544. Tyr985 recruits the tyrosine phosphatase PTPN11 [protein tyrosine phosphatase, non-receptor type 11 (also called SHP2) which controls extracellular-signal regulated kinase (ERK) activation] and also binds suppressor of cytokine signaling (SOCS) 3, an inhibitor of Lep Rb Jak2 signaling45.
B. IRS-PI3K pathway:
Intracellular signaling by insulin and leptin converge in key neuronal subsets at the level of the insulin receptor substrate-phosphatidylinositol-3-OH kinase (IRS-PI3K) pathway. A potential mechanism linking the leptin receptor to IRS-PI3K signaling involves the protein SH2B1 (46), which appears to facilitate Jak2-mediated IRS phosphorylation in response to leptin receptor activation Although leptin and insulin both have the potential to activate IRSPI3K signaling in neurons and other cell types, both the sub cellular localization and intracellular consequences of this activation can differ substantially depending on cell type and on whether activation is mediated by leptin or insulin. Leptin binding promotes the activation of LEPR-B-associated Jak2, which phosphorylates three tyrosine residues on the intracellular tail of LEPR-B (Fig. 3). Each of these phosphorylated residues recruits a unique set of downstream signaling molecules. Phosphorylated Tyr985 (pY985) recruits SHP2 (which participates in ERK activation) and SOCS3 (an inhibitor of LEPR-B signaling). pY1077 recruits the transcription factor STAT5, whereas pY1138 recruits STAT3. A variety of processes contributes to the attenuation of LEPR-B signaling (red lines), including the feedback inhibition that occurs by STAT3- promoted SOCS3 accumulation. PTP1B, ER stress and inflammatory signals might also participate in the inhibition of LEPR-B signaling in obesity.
CONCLUSION
It can be concluded from the various articles that signal from peripheral tissues, such as leptin hormone synthesized by adipose tissues, along with various other agents act at the central level to regulate energy intake, energy homeostasis and thereby controlling glucose and fat metabolism. The leptin hormone pathway may be a potential tool to fight against the rising menace of obesity and prevent the associated diseases like hypertension, myocardial infarction, and diabetes mellitus before their emergence.
ACKNOWLEDGEMENT
The authors acknowledge the immense help received from the scholars whose articles are cited and included in references of this manuscript. The authors are also grateful to authors / editors / publishers of all those articles, journals and books from where the literature for this article has been reviewed and discussed.
References:
1. Cherhab FF, Mounzih K, Lu R, Lim ME. Early onset of reproductive function in normal female mice treated with leptin. Science 1997, 275:88.
2. Gavrila A, Chan JL, Yiannakouris N, Kontogianni M, et al. Serum adiponectin levels are inversely associated with overall and central fat distribution but are not directly regulated by acute fasting or leptin administration in humans: Cross-sectional and interventional studies . J Clin Endocrinol Metab 2003 Oct;88 (10):4823-31.
3. Cnop M, Landchild MJ, Vidal J, Havel PJ, et al. The Concurrent Accumulation of Intra-Abdominal and Subcutaneous Fat Explains the Association Between Insulin Resistance and Plasma Leptin Concentrations Distinct Metabolic Effects of Two Fat Compartments. Diabetes 2002, 51:1005– 15.
4. Clement K, Vaisse C, Lahlou N, et al. A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature 1998, 392-398.
5. Buyse, M., Aparicio, T., Guilmeau, S., Goiot, H,et al. Paracrine actions of the stomach-derived leptin. Medecine Sciences 2004, 20:183-188.
6. Green ED., Maffei M. and Braden V.V. The human obese gene: RNA expression p pattern and mapping on the physical, cytogenetic and genetic of chromosome 7. Genome Research 1995, 5:5-12.
7. Smith-Kirwin, S.M., O’Connor, D.M., De Johnston, J.,Lancey, E.D., Hassink, S.G. and Funanage, V.L. Leptin expression in human mammary epithelial cells and breast milk. The Journal of clinical Endocrinology and Metabolism.1998; 83(5):1810-1813.
8. Friedman JM. and Halaas JL. Leptin and the regulation of body weight in mammals. Nature 1998; 395:763–770.
9. Elmquist JK, Maratos-Flier E., Saper CB. and Flier JS. Unraveling the central nervous system pathways underlying responses to leptin. Nature Neurosci. 1998; 1: 445–449.
10. Bates SH. and Myers MG Jr. The role of leptin receptor signaling in feeding and neuroendocrine function. Trends Endocrinol. Metab. 2003; 14: 447–452.
11. Considine RV, Sinha MK, Heiman ML, et al. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med. 1996; 334:292–5.
12. Frederich RC, Hamann A, Anderson S, Lollmann B, et al. Leptin levels reflect body lipid content in mice: evidence for diet-induced resistance to leptin action. Nat Med. 1995; 1:1311–4.
13. Halaas JL, Gajiwala KS, Maffei M, Cohen SL, et al. Weightreducing effects of the plasma protein encoded by the obese gene. Science 1995; 269: 543–6.
14. Faggioni R, Feingold KR and Grunfeld C. Leptin regulation of the immune response and the immuno- deficiency of malnutrition. FASEB J.2001; 15: 2565–71.
15. Cherhab FF, Mounzih K, Lu R and Lim ME. Early onset of reproductive function in normal female mice treated with leptin. Science 1997; 275:88-90.
16. Peter JH. Mechanisms regulating leptin production: implications for control of energy balance. Am J Clin Nutr. 1999; 70:305–6.
17. Hafeezullah, Muhammad A. Leptin: fights against obesity! Pak J Physiol 2006; 2(1):1-7.
18. Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature 1998; 395:763-75.
19. Cunningham MJ, Clifton DK and Steiner RA. Leptin’s actions on the reproductive axis: perspectives and mechanisms. Biol Reprod 1999; 60: 216–22.
20. Loffreda S, Yang SQ, Lin HZ, et al. Leptin regulates proinflammatory immune responses. FASEB J 1998; 12:57–65.
21. O’Donnell CP, Schaub CD, Haines AS, et al. Leptin prevents respiratory depression in obesity. Am J Respir Crit Care Med 1999; 159: 1477–84.
22. Schwartz MW, Baskin DG, Kaiyala KJ, Woods SC. Model for the regulation of energy balance and adiposity by the central nervous system. Am J Clin Nutr 1999; 69:584–96.
23. Barzilai N, Wang J, Massilon D, Vuguin P, et al. Leptin selectively decreases visceral adiposity and enhances insulin action. J Clin Invest 1997; 100: 3105–10.
24. Bates SH. and Myers MG Jr. The role of leptin receptor signaling in feeding and neuroendocrine function. Trends Endocrinol. Metab. 2003; 14: 447–452.
25. Ahima RS, Prabakaran D, Mantzoros CS, Qu D, et al. Role of leptin in the neuroendocrine response to fasting. Nature 1996; 382: 250–252.
26. Gregory J. Morton and Michael W. Schwartz. Physiol Rev. 2011 April ; 91(2): 389–411.
27. Tartaglia LA. The leptin receptor. J. Biol. Chem. 1997; 272:6093–96.
28. Chua SC Jr, Koutras IK, Han L, Liu SM, et al. Fine structure of the murineleptin receptor gene: Splice site suppression is required to form two alternatively spliced transcripts. Genomics 1997; 45:264–70.
29. Bates SH, Myers MG Jr. The role of leptin receptor signaling in feeding and neuroendocrine function. Trends Endocrinol. Metab. 2003; 14:447–52.
30. Elmquist JK, Elias CF, Saper CB. From lesions to leptin: hypothalamic control of food intake and body weight. Neuron 1999; 22:221–32.
31. Elmquist JK, Bjorbaek C, Ahima RS, Flier JS, et al. Distributions of leptin receptor mRNA isoforms in the rat brain. J. Comp. Neurol 1998; 395:535–47.
32. Baskin DG, Schwartz MW, Seeley RJ, Woods SC, et al. Leptin receptor long-form splice-variant protein expression in neuron cell bodies of the brain and colocalization with neuropeptide Y mRNA in the arcuate nucleus. J. Histochem. Cytochem. 1999; 47:353–62.
33. Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, et al. Central nervous system control of food intake. Nature 2000; 404:661–71.
34. Huo L, Munzberg H, Nillni EA, Bjoraek C. Role of signal transducer and activator of transcription 3 in regulation of hypothalamic trh gene expression by leptin. Endocrinology 2004; 145:2516–23.
35. Elmquist JK, Ahima RS, Maratos-Flier E, Flier JS, et al. Leptin activates neurons in ventrobasal hypothalamus and brainstem. Endocrinology 1997; 138:839–42.
36. Martin GM, Michael AC, and Heike M. Mechanisms of Leptin Action and Leptin Resistance. Annu. Rev. Physiol. 2008; 70:537-556.
37. Liu L, Karkanias GB, Morales JC, Hawkins M, et al. Intracerebroventricular leptin regulates hepatic but not peripheral glucose fluxes. J. Biol. Chem.1998; 273: 31160–31167.
38. Kulkarni RN, Wang ZL, Wang RM, Hurley JD, et al. Leptin rapidly suppresses insulin release from insulinoma cells, rat and human islets and, in vivo, in mice. J. Clin. Invest. 1997; 100: 2729–2736.
39. Burcelin R, Kamohara S, Li J, Tannenbaum GS, et al. Acute intravenuous leptin infusion increases glucose turnover but not skeletal muscle glucose uptake in ob/ob mice. Diabetes 1999; 48: 1264–1269.
40. Kieffer TJ, Heller RS, Leech CA, Holz GG, et al. Leptin suppression of insulin secretion by the activation of ATP-sensitive K+ channels in pancreatic betacells. Diabetes 1997; 46: 1087–1093.
41. Robertson S.A, Leinninger G.M. and Myers M.G Jr. et al. Molecular and neural mediators of leptin action. Physiol. Behav 2008; 94: 637–642.
42. Gong Y, Ishida-TR, Villanueva EC, Fingar DC, et al. The long form of the leptin receptor regulates STAT5 and ribosomal protein S6 via alternate mechanisms. J. Biol. Chem. 2007; 282: 31019–31027.
43. Munzberg H, Flier JS, Bjorbaek C, et al. Region-specific leptin resistance within the hypothalamus of diet-inducedobese mice. Endocrinology 2004; 145: 4880–4889.
44. Hekerman P, Zeidler J, Bamberg-Lemper S, et al. Pleiotropy of leptin receptor signalling is defined by distinct roles of the intracellular tyrosines. FEBS J 2005; 272: 109–119.
45. Martin GM Jr, Rudolph L. Leibel, Randy JS, et al. Obesity and leptin resistance: distinguishing cause from effect. Trends in Endocrinology and Metabolism 2011; 21(11): 643-651.
46. Li M, Li Z, Morris DL, Rui L. Identification of SH2B2beta as an inhibitor for SH2B1- and SH2B2alpha-promoted Janus kinase-2 activation and insulin signaling. Endocrinology 2007; 148:1615–1621.
|
 IJCRR
IJCRR 





 This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License





