IJCRR - 4(7), April, 2012
Pages: 25-32
Print Article
Download XML Download PDF
PRENATAL STUDY OF HISTOGENESIS OF HUMAN LATERAL GENICULATE BODY
Author: Indira Devi B, Raju Sugavasi, Sujatha M, Sirisha B
Category: Healthcare
Abstract:Objectives: The lateral geniculate body in human fetuses of different gestations. The Present study is to determine the apperance of the number of laminae and to observe the types of cells present in different laminae at different gestations. Methods: The present work is the result of study of lateral geniculate bodies in human fetuses, which are obtained from local government hospitals. Age of the fetuses ranged from 20-40 weeks. The age of fetuses is judged by CR lengths. The photo graphes of slides are taken by SAMTRON'computer having a closed circuit camera and an adapter fixed to ?LYNX' (LAWRENCE AND MAYO) trinocular research microscope after the routine histological procedures. Result: Histo genisis of lateral geniculate body is of different propotions at different gestations of embryonic life. In the present study at 20 wees of gestation bipolar cells are abundantly seen and laminar differentiation has not seen. At 24 th week of geststion the laminar differentiation has started.all 6 laminae are visible by at 33 weeks of gestations.varied diameters of rounded neurons are seen with predominance of multipolar neurons have been observed at 38 weeks of gestation. Multipolar cells are visible at 40 weeks of gestation in all 6 laminae. Conclusion: in the present study all 6 laminae are differentiated at 33 week of gestation. During 20 to 40 weeks of gestation the predominance of cells present in laminae differentiated from rounded to multipolar neurons.
Full Text:
INTRODUCTION
In view of, interesting changes that occur in the cytoarchitecture of lateral geniculate body, an attempt has been made to notify its histogenesis in human fetuses of different gestations. The lateral geniculate body is a part of the visual pathway. It is a small, ovoid, ventral projection from the posterior thalamus. The lateral geniculate nucleus is an inverted, somewhat flattened, U-shaped nucleus and is laminated. Its internal organization is usually described on the basis of six laminae, although seven or eight may be present. The laminae are numbered 1 to 6, from the innermost ventral to the outermost dorsal. Laminae 1 and 2 consist of large cells, the magnocellular layers, whereas layers 4 to 6 have smaller neurones, the parvocellular laminae. The apparent gaps between laminae are called the interlaminar zones.
The lateral geniculate nucleus receives major afferent inputs from the retina. The contralateral nasal hemiretina projects to laminae 1, 4 and 6, whereas the ipsilateral temporal hemiretina projects to laminae 2, 3 and 5. The efferent fibres from the lateral geniculate nucleus pass principally to the primary visual cortex (area 17) in the banks of the calcarine sulcus. It is possible that additional small projections from interlaminar zones pass to extrastriate visual areas in the occipital lobe [1] .
MATERIALS AND METHODS
The present work is the result of study of lateral geniculate bodies in human fetuses. 50 human fetuses have taken up and all the fetuses belong to gestational age group of 20 – 40 weeks. The age of fetuses is judged by CR lengths. The foetal brains have been fixed by injecting 10% formalin into the vessels of umbilical cord and through anterior fontanelle. The cranial vaults have been removed by using a bone cutter, hand saw and scissors. The dural folds have been removed to expose the cerebral hemispheres. The brains have been severed at the lower end of medulla oblongata. It has been observed that most of foetal brains have not hardened, most probably foetal brains are having less neuroglial tissue.The piamater has been removed. The optic tracts are traced backwards to view the lateral geniculate bodies. After the fixation, tissue processing, and sectioning the microphotographs of sections stained with haematoxylin and eosin were studied on a ?SAMTRON‘ computer having a closed circuit camera and an adapter fixed to ?LYNX‘ (LAWRENCE AND MAYO) trinocular research microscope. The stained sections were examined with 10x and 40x magnifications. The closed circuit camera with adapter is attached to one of the eye pieces of trinocular microscope. With manipulation of the fine adjustment of camera as well as that of microscope, pictures having magnifications of 10x and 40x have been obtained with good resolution on computer screen and this has been utilized for taking microphotographs of various sections. The lateral geniculate body in human fetuses of different gestations is further studied, to view the number of laminae and observe the types of cells present in different laminae.
RESULTS
Microscopic Observations:
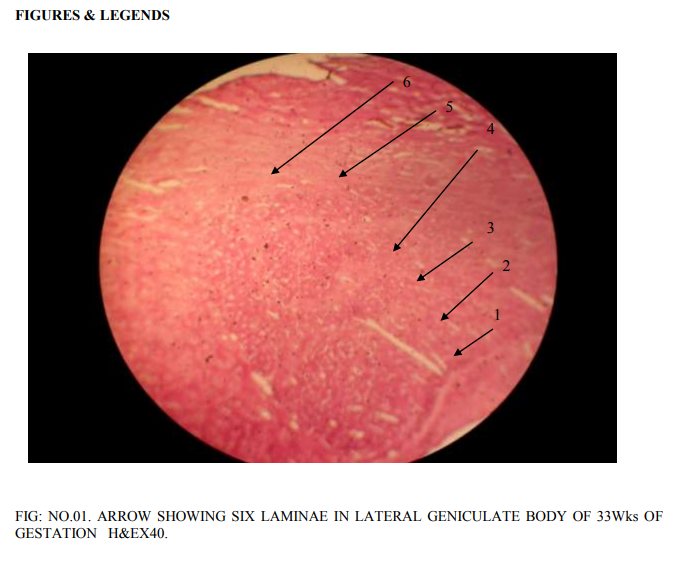
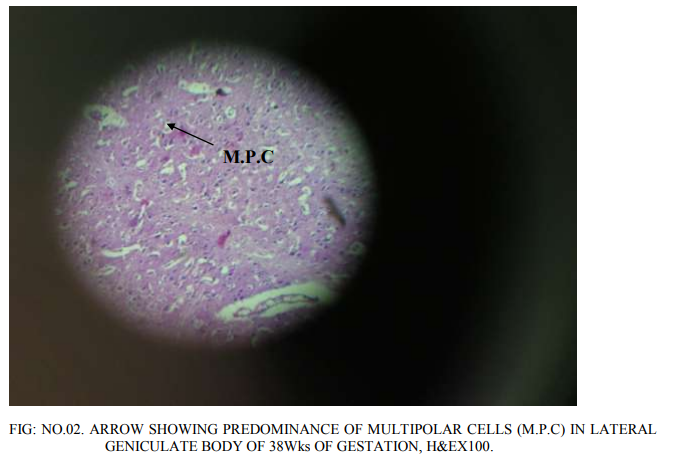
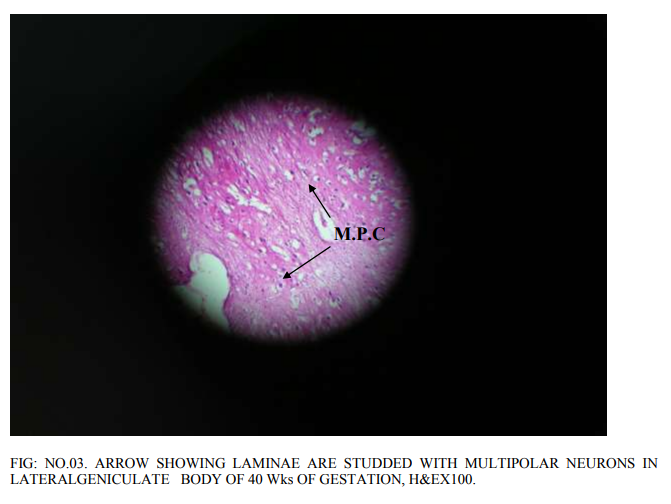
Lateral geniculate body is of different proportions at different gestations of the embryonic life. Most of the neurons in it are rounded to oval to multi polar. The density of the neurons as well as congregation of neurons changed as the gestations of fetuses increases. Before 20th week, bipolar cells are abundant. However, a few multi polar cells are also seen. 20 weeks of gestation: The proportion of bipolar and multi polar cells is 4:1. No clear laminar pattern is seen. There is aggregation of the rounded neurons into nuclear groups. However, definite nuclear groups are not clearly seen. Division of magnocellular and parvocellular components of lateral geniculate body is also not clear. Supporting glial cells are abundant. 24 weeks of gestation: Laminar pattern is still not clear, Neuron density is distinctly more. The number of multi polar neurons has increased. Differentiation of parvocellular and magnocellular regions has begun. Axodendritic synapsis started appearing. There is increased density of glial cells simultaneously. Increased vascularity is also seen, confirming the increased cellular activity. 33 weeks of gestation: 6 laminae are visible (Fig.no.01). Each lamina is separated by a white band that indicates the synaptic zone in between the cells of the different lamina. The first lamina is concave and cells are small, multi polar cells, the density of the cells is less. The second lamina is having higher density of multipolar cells. The cell nuclei are closely packed. In the synaptic zone round to oval cells are present, these are called koniocytes or konio (k) cells. Third lamina is thicker than that of the first and second lamina. Cells are widely distributed throughout entire length of the lamina. Most of the cells are still rounded. The density of multi polar cells is less. The glial cells are also more. The width of the fourth lamina is less than that of the third lamina. Cells are widely distributed. Cell nuclei are small and rounded. Multi polar cells are also seen, but minimal in number.The fifth and sixth lamina are clearly demarcated like the rest of the laminae. The cells of the fifth lamina are multipolar and rounded. Similarly, in the sixth lamina also the cells are multipolar and rounded. 38 weeks of gestation: A well developed 6 layerd lateral geniculate body with predominance of multipolar cells of varied diameter with very minor density of rounded neurons has been observed (Fig.no.02).The conversion of rounded neurons into multipolar neurons will be taking place henceforth. Koniocytes are clearly seen in between the laminae. 40 weeks of gestation: All the 6 laminae are clearly seen.Each of the lamina is studded with multipolar neurons (Fig.no.03). Koniocytes are also seen in between the laminae. No increase in the width of the lamina is seen.
DISCUSSION
Lateral geniculate nucleus is a thalamic station in the projection of visual pathway from retina to primary visual cortex. The topographic organization and the study of neurons have been extensively taken up in non human primates by Polyak (1953) [2], Kaas et al (1972) [3], Malpeli and Baker (1975) [4]. The lateral geniculate body is organized into six layers, each of which receive input from either the contralateral or ipsilateral eye and contains a retinotopic map of lateral hemifield. Out of six layers, the dorsal layers contain small neurons where as the ventral two layers contain large neurons. Hence, the distinction of the two regions of dorsal parvocellular and ventral magnocellular components has been observed by Wiesel and Hubel (1966) [5], Creutzfeldt et al (1979) [6], Maunsell (1993)[7] . The development of the human lateral geniculate body is remarkably similar to that of Rhesus monkey (Rakic, 1977) [8]. The lateral geniculate body is displaced and rotated infero lateral to the thalamus because of late growth of pulvinar around the 24`Th week of intra uterine life and the lateral geniculate body has come to lie along the ventro lateral border of the thalamus. The characteristic laminar pattern has been observed over a period of 3 weeks, beginning around 22 weeks of gestation. Caudal part of the lateral geniculate body laminates first (Hickey and Guillery 1979) [9] . In the earlier studies, it was found out that the lateral geniculate body became apparent at 14weeks, light microscopically (Dhamayanti N, S. Wadhwa and V. Bijlani, 1988) [10].At 16 weeks, the nucleus attained the maximum size and was found to be having two parts, the larger part closely associated with sub thalamus another less distinct part among the optic tract fibres. At 18 to 20 weeks, the appearance of the lateral geniculate nucleus is similar as at 16 weeks.At 22 weeks the nucleus was situated rostrally. Saini K, Kretz R and Rager G (1987) [11] described three types of neurons, these are multipolar radiate, bi tufted and intermediate types. The dendrites of these neurons are confined to one lamina only. Multi laminar neurons with multipolar radiate, bi tufted and intermediate types of dendrites have also been described by them. Inter laminar neurons whose cell bodies and dendrites are confined to a single inter laminar zone have also been described. The lateral geniculate nucleus (LGN) is the principal thalamic relay to the visual cortex (area 17), and its neurons have similar morphological characteristics in both monkey and man, as identified by Golgi impregnation (de Courten C, Garey LJ, 1983) [12]. Hitchcock PF, Hickey TL (1980) [13] stated that the development of the human lateral geniculate nucleus is remarkably similar to that described for the rhesus monkey (Rakic, 1977). The development of the lateral geniculate nucleus has been studied systematically in Nissl preparations from a series of cats (Kalil R) [14] that ranged in age from newborn to adult. In addition, preliminary observations are reported at two stages of fetal development. It was found that laminae develop in the lateral geniculate nucleus near the time of birth and continue to differentiate during the first postnatal week. During development the major axis of the lateral geniculate rotates approximately 180 degrees in the sagittal plane. The rotation begins prenatally and is not completed until after the twentieth postnatal week. In the human lateral geniculate nucleus (LGN) anatomical studies have revealed a similar organisation compared with the macaque in lateral geniculate nucleus (LGN) terms of laminar pattern. The layout of the representation of the visual field, however, is less well understood, because its study has historically been restricted to postmortem anatomical analysis of degeneration patterns after retinal or cortical lesions (Rönne, 1910 [15] , Juba and Szatmári, 1937 [16], Highresolution functional magnetic resonance imaging (fMRI) at 3 T in humans has been utilized to derive a detailed account of the retinotopic organisation of the lateral geniculate nucleus (LGN). The results of researchers reveal a close correspondence in the topographic organisation of the macaque and human lateral geniculate nucleus (LGN) and support accounts of a constant magnification from the retina through the cortex.
CONCLUSION
In the present prenatal histological study At 20 weeks gestation bipolar cells are abundantly seen. Formation of nuclear groups is observed. Division of magnocellular and parvo cellular components has not taken place. Differentiation of parvocellular and magnocellular regions has started at 24 weeks there is increased density of neurons and glial cells. All 6 layers are visible by 33 weeks. The first lamina is concave and is present towards the hilum. The first and second laminae of the lateral geniculate body have closely packed with multipolar cells at 33 weeks of gestation. The density of glial cells is more in 3`rd lamina with round cell nuclei. The 5`Th and 6`Th laminae are not clearly distinguished, with predominance of multipolar neurons. Varied diameters of rounded neurons are seen with predominance of multipolar neurons is observed at 38 weeks of gestation. All 6 layers are well developed. All the neurons are of 6 laminae are multipolar at 40 weeks of gestation.
ACKNOWLEDGEMENTS
I express my deep sense of gratitude to Dr. B. Narasinga Rao professor and HOD of Anatomy and Padmini for well support through out this work, and colleagues for their proper guidance, precious suggestions and priceless encouragements to accomplish my work. I am very much greatful to the research scholars and so many authors whose efforts have helped me to update my knowledge of Anatomy and continue this work. I am thankful to my co authors for contribution of their efforts to this work.
References:
1. Standring S. Grays Anatomy. The Anatomical basis of clinical practice. 40 Th ed. Edinburg. Elsevier Churchill Livingstone ; 2008. (21): p. 314 - 315.
2. Polyak S, Santiago Ramon y Cajal and his investigation of the nervous system. J Comp Neurol. 1953: 98: 3 - 8.
3. Kaas JH, Guillery WR, Allman JM, Some principles of organization in the dorsal lateral geniculate nucleus. Brain Behav Evol. 1972; 6: 253 - 299.
4. Malpeli JG, Baker FH. The representation of the visual field in the lateralgeniculate nucleus of Macaca mulatta. J Comp Neurol. 1975: 161: 569 - 594
5. Wiesel TN, Hubel DH. Spatial and chromatic interactions in the lateral geniculate body of the rhesus monkey. J Neurophysiol. 1966: 29: 1115 - 1156.
6. Creutzfeldt OD, Lee BB, Elepfandt A. A quantitative study of chromatic organisation and receptive fields of cells in the lateral geniculate body of the rhesus monkey. Exp Brain Res. 1979: 35: 527 - 545.
7. Maunsell JH, Merigan WH. How parallel are the primate visual pathways? Annu Rev Neurosci. 1993: 16: 369 - 402.
8. Rakic P. Prenatal development of visual system in rhesus monkey, Phil Trans R Soc Lond, 1977; B278:245.
9. Hickey TL, Guillery RW. Variability of laminar patterns in the human lateral geniculate nucleus. J Comp Neurol. 1979: 183: 221 - 246.
10. Bijlani V, Wadhwa S. Cytodifferentiation and developing neuronal circuitry in the human lateral geniculate nucleus. Int J Dev Neurosci. 1988; 6 (1): 59 - 75.
11. Saini K, Kretz R, Rager G. Classes of neurons in relation to the laminar organization of the lateral geniculate nucleus in the tree shrew, Tupaia belangeri. J Comp Neurol. 1987 May 1;259(1):31 – 49.
12. de Courten C, Garey LJ. Morphology of the neurons in the human lateral geniculate nucleus and their normal development. A Golgi study.Exp Brain Res. 1982 ;47(2):159 - 71.
13. Hitchcock PF, Hickey TL. Prenatal development of the human lateral geniculate nucleus, J Comp Neurol. 1980 Nov 15; 194(2): 395 - 411.
14. .Kalil R. Development of the dorsal lateral geniculate nucleus in the cat. J Comp Neurol 1978:No15; 182(2): 265 - 91.
15. RonneH Pathologisch-anatomische untersuchungen über alkoloische intoxikationsamblyopie. Arch Ophthalmol. 1910: 77: 1 - 95.
16. Juba A, Szatmári A. Ueber seltene hirnanatomische befunde in fällen von einseitiger peripherer blindheit. Klin Monatsbl Augenheilkd. 1937: 99: 173 - 188.
|
 IJCRR
IJCRR 





 This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License





