IJCRR - 4(16), August, 2012
Pages: 39-46
Date of Publication: 28-Aug-2012
Print Article
Download XML Download PDF
ANTAGONISTIC ACTIVITY OF MARINE BACILLUS SPECIES FROM MARINE SEDIMENT OF PUNNAIKAYAL ESTUARY OF TUTICORIN AGAINST ASPERGILLUS NIGER
Author: V.Bharathi, Jamila Patterson, R. Rajendiran
Category: General Sciences
Abstract:A total of 15 marine Bacillus species were isolated from estuarine sediment of Punnaikayal estuary of Tuticorin, Tamilnadu, India. The marine Bacillus was evaluated for biocontrol activities against Aspergillus niger a phytopathogenic fungus causing Aspergillus rot of papaya, collar rot of groundnut, post harvest disease, food spoilage and human pathogen. In vitro dual culture and volatile assay of marine Bacillus and A. niger on agar plates showed that the bacterium effectively inhibited radial growth of the fungus. Among the inhibitory marine Bacillus isolates, MB-13 and MB-6 significantly suppressed hyphal growth of A. niger in dual culture assay and MB-6, MB-13, MB-5 isolates inhibit the growth of A. niger by volatile assay. These findings support the potential use of marine Bacillus also for biological control of plant pathogens.
Keywords: Biological Control, Dual culture assay, Volatile assay, Black mould, Marine Bacillus species.
Full Text:
INTRODUCTION
Diseases of plants by fungi can cause vast crop damages which may lead to hunger disasters of great part of populations. Different methods may be used to prevent, mitigate or control plant diseases. Beyond good agronomic and horticultural practices, producers often rely heavily on chemical fertilizers and pesticides (Pal and McSpadden Gardener, 2006). Synthetic chemicals have been used to control plant diseases, but abuse in their employment has favored the development of pathogens resistant to fungicides (Benitez et al., 2004). Chemical compounds have been used to control plant diseases, but abuse in their application has favored the development of pathogens resistant to fungicides (Yaqub and S.Shahzad, 2009). By contrast, the potential use of microbes based biocontrol agents that antagonize plant pathogens is slow, gives few quick profits, but can be long lasting, inexpensive and harmless to life (Zamanian et al., 2005). During the past decades, various groups of microorganisms including bacteria, yeasts and filamentous fungi have been screened for antagonistic activities against certain plant pathogens and numerous effective antagonists have been reported (Siripornvisal, 2010). Among them, Bacillus species such as B. subtilis (Xiaoxi and Liu Wenhong, 2011), B. licheniformis (Kong et al., 2010), B. megaterium, B. cereus, B. pumilus, and B. polymyxa (Utkhede, 1984; Silo et al., 1994) are the most promising antagonists due to its ability to produce various antimicrobial compounds which contribute to its biocontrol capability. The genus Bacillus, or use of their metabolites, may be an alternative or supplementary method to chemical plant protection to control of plant disease (Leifert et al., 1995; Berger et al., 1996; Sharga and Lyon 1998). The aquatic environment, representing more than two thirds of our planet, is still underexplored and is considered to be a prolific resource for the isolation of less exploited microorganisms (Sponga et al., 1999) and to produce new antifungal substances. Preliminary data was encouraging marine isolates produce antibiotic activities with frequencies comparable to terrestrial ones (Burgess et al., 1999). Most of recent studies on marine microorganisms are more studied on human drugs than plant pathogens. Therefore the objective of this investigation was to explore the potential of biological control of plant pathogen, Aspergillus niger with marine Bacillus species in vitro condition.
MATERIALS AND METHODS
Isolation of marine Bacillus isolates
Estuarine sediment was collected from Punnaikayal estuarine environment of Tuticorin on July 2011 and brought to laboratory in polythene bag. For isolation of Bacillus species, Each 1g of the sample was suspended in 9 ml of half strength sterile sea water and shaken vigorously for 2 min. The samples were heated at 80oC for 40 min in a water bath (Al-Ajlani and Hasnain, 2010). The suspensions were serially diluted and spread on nutrient agar medium. The plates were incubated at 28 - 37° C for 24 - 48 h. Individual colonies were isolated on the basis of their different visual characteristics and identified as Bacillus species by grams and spore staining. Test strain: A. niger was used as test strain which isolated from market waste of Tutucorin District. The strain of A. niger was maintained on slant tubes of PDA medium at 4°C and cultivated on plates of PDA medium at 23°C for at least 4 days for sufficient growth of conidia. The conidial suspension of A. niger was prepared by adding the conidia to a sterile saline solution (0.85% NaCl), counting the conidia with a haemocytometer and adjusting to 1x104 conidia/ml. Antagonistic activity of Marine Bacillus isolates against A. niger
1) Dual culture method
For detection of antagonistic activity of bacterial isolates towards the growth of the fungal cultures Montealegre et al., (2003) methods were used. One 10-mm disk of a pure culture of A. niger was placed at the center of a Petri dish containing PDA. All the plates were individually and equally marked into two parts. In the peripheral region of a part of a plate was streaked with the Bacillus isolates (5 x 109 cfu mL1), the second part of the peripheral region just three cm away from the edge of the plates, but direct opposite to first part of peripheral region was inoculated by using a 5mm mycelial disc of A. niger. The same procedure was followed to all the plates. In the control experiment an A. niger disc only placed on agar plate without bacterial streaking. Plates were cultured for 72 hours at 22ºC and growth diameter of the pathogen was measured and compared to control growth where the bacterial suspension was replaced by sterile distilled water. Each experiment considering a single A. niger isolate was run in triplicate and was repeated at least three times. Results are expressed as means % inhibition of the growth of the corresponding A. niger isolate in the presence of any of the bacterial isolate. The percentage inhibition of the fungus was calculated with the following formula (Sivakumar et al., 2000)
C – T X 100 C Where, C is radial growth measurement of the pathogen in control and T is radial growth of the pathogen in the presence of marine Bacillus isolates.
2) Volatile antagonistic activity
The volatile antagonistic activity described by Dennis and Webster, (1971) was used for the in-vitro test of antagonistic activities of bacterial isolates against test fungi. Bacillus isolates were grown on nutrient agar plates (90mm) at 28±2?C. After 16 - 20hours, the lid of the plate was replaced by a bottom plate containing 20ml of PDA and spot inoculated with test fungus. The two plates were sealed together with adhesive transparent cello – tape. Plate without Bacillus isolates served as control at 28±2?C. After 48 hours, the percentage of growth inhibition of the test fungus was calculated with the following formula r1 – r2 X 100 r1 Where, r1 is radial growth of a fungus in control sets (i.e. without the bacterium) and r2 is the radial growth of a fungus in sets inoculated with the bacterium.
Satistical analysis
The statistical software package Minitab version 15 (Minitab Ltd., Coventry CV3 2TE, UK) was used for analyzing the experimental data.
RESULTS AND DISCUSSION
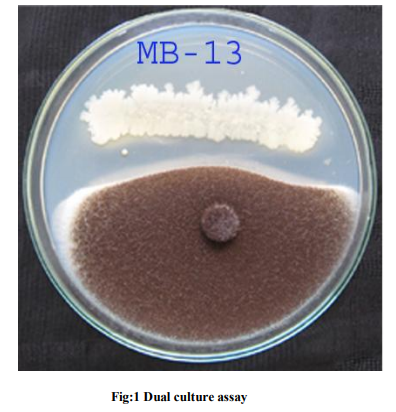
Aspergillus niger Van Teighem cause Aspergillus rot of papaya (Llondu, 2011), collar rot of groundnut (Chohan, 1969), post harvest disease (Ogawa et al., 1995), food spoilage (Suman Singh, 2010) and human pathogen (Bansod and Rai, 2008). This universal fungus can be controlled by antagonistic microorganisms. Many of the genus Bacillus and their secondary metabolites have been found to show antibacterial and/or antifungal activity against phytopathogenic and food borne pathogenic microorganisms (Moyne et al., 2001). Due to the ability to form endospores, Bacillus species exist in many various environments (Leifert et al., 1995; Foldes et al., 2000). The fifteen isolates come from the marine environment which is morphologically similar with terrestrial habitats. Its potential as a biocontrol agent for black mould is interesting and worth for further investigation. Bacillus isolates effectively inhibited the mycelia growth of A.niger, A.fumigatus, A.flavus, A.tereus, A.glacus (Bottone and Peluso, 2003) by production of secondary metabolites (Krebs et al., 1996). Table 1 and figure 1 showed the MB13 isolate showed maximum inhibitory activity (53.16%) on radial growth of A. niger on 96 hours incubation followed by MB6. The isolates like MB1, MB5, MB8, MB9, MB10 and MB11 showed moderate inhibitory action against A. niger. In general, volatile compound from Bacillus sp. have been investigated for different purposes and the compounds have been used as inhibitors of fungi and bacteria (French, 1992).
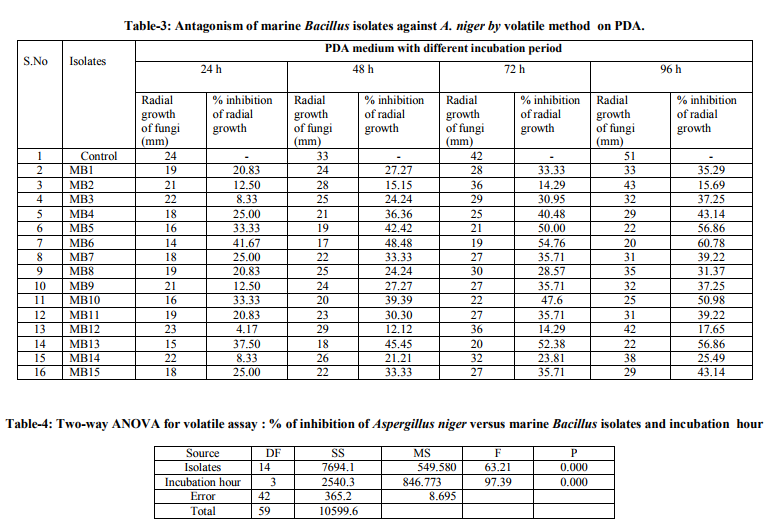
In table 3, maximum percent inhibition of radial growth of fungi was observed with isolates of MB6-1 (60.78%) followed by MB 5 and MB13 (56.86%) and MB10 (50.98%). The remaining isolates were inhibited the radial growth of A. niger in the range of medium to low. In table 2 and 4 represented two way ANOVA of dual culture assay and volatile assay respectively, which clearly indicated to extend of inhibition increased with increase in incubation period. Bacillus isolates produced metabolites like bacitracin, bacillin, bacillomycin B, bacilysin, which belong to iturin family (Parry et al., 1983), phenazine (Pierson and Thomashow, 1992) and antibiotics like difficidin and oxdifficidin (Zimmerman et al., 1987) antifungal peptides such as iturin (Krebs et al., 1996) highly influenced to inhibit the growth of A. niger. The variation of A. niger inhibition may have been due to the fungus may respond to different strains of Bacillus, or that variation in antifungal activity may reflect differences in the site of action or the ability of fungi to detoxify the metabolites. It may have also been due to the inactivation of the antibiotics in the environment.
CONCLUSION
Marine Bacillus isolates MB-6 and MB13 considered as best isolate which significantly diminished the growth of A. niger. From all these results it may be concluded that the biocontrol effect of antagonistic bacteria isolated from estuarine source against A. niger are adequate for their use at the various field. Within the mechanisms used by these bacteria are the secretions of volatile and difusible metabolites and also fungal cell wall hydrolytic enzymes. Therefore, these bacteria could be used for biocontrol the A. niger disease in the agriculture, food spoilage and post harvest disease. Even more investigation is needed to elucidate the modes of action of marine Bacillus species against A. niger and other fungal pathogens and the condition in which the biocontrol isolate can be used effectively.
ACKNOWLEDGEMENT
Authors are thankful to the Director of Suganthi Devadason Marine Research Institute, Tuticorin, TamilNadu, India for permitting to explore the field of biological control of plant pathogenic fungi using marine Bacillus species. We are also thankful to the Poultech Agro Research centre Bioinput Production Unit, Namakkal, TamilNadu, India for permitting to do the research work
References:
1. Al-Ajlani MM, Hasnain S. Bacteria Exhibiting Antimicrobial Activities; Screening for Antibiotics and the Associated Genetic Studies. The Open Conference Proceedings Journal 2010; 1: 230-8.
2. Bansod S, Rai M. Antifungal activity of essential oils from indian medicinal plants against human pathogenic Aspergillus fumigatus and A. niger. World J Med Sci 2008;3: 81-88.
3. Benitez T, Rincon AM, Carmen Limon MC, Codon AC. Biocontrol mechanisms of Trichoderma strains. Inter J Microbiol 2004; 7(4): 249-60.
4. Berger F, Li H, White D, Frazer R, Leifert C. Effect of pathogen inoculum, antagonist density, and plant species on biological control of Phytophtora and Pythium damping-off by Bacillus subtilis Cot1 in high-humudity fogging glasshouses. Phytopathology 1996; 86:428-433.
5. Bottone EJ, Peluso RW. Production by Bacillus pumilus (MSH) of an antifungal compound that is active against Mucoraceae and Aspergillusspecies: preliminary report. J Med Microbiol 2003;52:69-74.
6. Burgess JG, Jordan EM, Bregu M, MearnsSpragg A, Boyd KG. Microbial antagonism: a neglected avenue of natural products research. J Biotechnol 1999; 70:27-32.
7. Chohan JS. Survival of Aspergillus niger V. Teighem and soil factors influencing collar rot disease of ground nut. J Res 1969; 6:634-641.
8. Dennis C, Webster J. Antagonistic properties of species groups of Trichoderma 11. Production of volatile antibiotics.Trans Br Mycol Soc 1971; 57: 41-48.
9. Foldes T, Banhegyi T, Herpai Z, Varga L, Szigeti J. Isolation of Bacillus strains from the rhizosphere of cereals and in vitro screening for antagonism against phytopathogenic and spoilage microorganisms. J Appl Microbiol 2000; 89: 840 -845.
10. French R C. Volatile chemical germination stimulators of rust and other fungal spores. Mycologia 1992; 84:277-288.
11. Ilondu EM. Evaluation of some aqueous plant extracts used in the control of pawpaw fruit (Carica papaya L.) rot fungi. J Appl Biosci 2011; 37: 2419 – 2424.
12. Kong HG, Kim JC, Choi GJ, Lee KY, Kim HJ, Hwang EC, et al. Production of Surfactin and Iturin by Bacillus licheniformis N1 Responsible for Plant Disease Control Activity. Plant Pathol J 2010; 26(2) : 170-177.
13. Krebs CJ, Sinclair ARE, Boutin S. Vertebrate community dynamics in the boreal forest of North-western Canada. In Frontiers and Applications of Population Ecology, ed. by R.B. Floyd, A.W. Sheppard, and P.J. De Barro, CSIRO Publishing, Collingwood, Victoria, Australia 1996; 155-161.
14. Leifert C, Workman S, Li H. Antibiotic production and biocontrol activity by Bacillus subtilis CL27 and Bacillus pumilus CL45. J Appl Bacteriol 1995; 78: 97-108.
15. Montealegre RJ, Reyes R, Perez ML, Herrera R, Silva P, Besoain X. Selection of bioantagonistic bacteria to be used in biological control of Rhizoctonia solani in tomato. Electron J Biotechnol 2003; 6:115-27.
16. Moyne AL, Shelby R, Cleveland TE, Tuzum S. Bacillomycin D: an iturin withantifungal activity against Aspergillus flavus. J Appl Microbiol 2001; 90:622-29.
17. Ogawa JM, Dehr EI, Bird GW, Ritchie DF, Kiyoto V, Uyemoto J.K. Compendium of Stone fruit Diseases. 1995; APS Press, US.
18. Pal KK, McSpadden Gardener B.Biological Control of Plant Pathogens. The Plant Health Instructor DOI 2006; 10:1094/PHI-A-2006- 1117-02.
19. Parry JM, Turnbull PCB, Gibson JR. A Colour Atlas of Bacillus Species. 1983; Wolfe Medical Publications, Ltd., London.
20. Pierson LS, Thomashow LS. Cloning and heterologous expression of the phenazine biosynthetic locus from Pseudomonas aureofaciens 30-84. Mol Plant-Microbe Interact 1992; 5:330-39.
21. Sharga BM, Lyon GD. Bacillus subtilis BS 107 as an antagonist of potato blackleg and soft rot bacteria. Can J Microbiol 1998; 44:777-83.
22. Silo-suh LA, Lethbridge BJ, Raffel SJ, He H, Clardy J, Handesman J. Biological activities of two fungistatic antibiotics produced by Bacillus cereus UW85. Appl Environ Microbiol 1994; 60, 2023-30.
23. Siripornvisal S. Biocontrol Efficacy of Bacillus subtilis BCB3-19 against Tomato Gray Mold KMITL Sci Tech J 2010; 10(2): 45-51.
24. Sivakumar D, Wilson RS, Wijesundera RLC, Marikar FMT, Abeyesekere M. Antagonistic effect of Trichoderma on postharvest pathogens of Rambutan (Nephelium lappaceum). Phytoparasitica 2000; 28:240-47.
25. Sponga F, Cavaletti L, Lazzarini A, Borghi A, Ciciliato I, Losi D, Marinelli F. Biodiversity and potentials of marine-derived microorganisms. J Biotechnol 1999; 70:65-9.
26. Suman Singh. In vitro antifungal activity of some essential oils against food spoilage fungi. J Herbal MedToxicol 2010;4(2):107-11.
27. Utkhede RS. Antagonism of isolates of Bacillus subtilis to Phytophthora cactorum. Can Bot 1984; 62: 1032-35.
28. Xiaoxi and Liu Wenhong, Potent antagonistic activity of newly isolated biological control Bacillus subtilis and novel antibiotic against Erysiphe graminis f.sp tritici. Chen- J Med Plants Res 2011; 5(10): 2011-14.
29. Yaqub F, Shahzad S. Effect of solar heating by polyethylene mulching on sclerotial viability and pathogenicity of Sclerotium rolfsii on mungbean and sunflower. Pak J Bot 2009; 41(6): 3199-205.
30. Zamanian S, Shahidi Bonjar GH, Saadoun I. First report of antibacterial properties of a new strain of Streptomyces plicatus (strain 101) against Erwinia carotovora from Iran. Biotechnology 2005; 4:114-120.
31. Zimmerman SB, Schwartz CD, Monaghan RL, Pelak BA, Weissberger B, Gilfillan EC. et al. 1987. Difficidin and oxydifficidin: novel broad spectrum antibacterial antibiotics produced by Bacillus subtilis. I. Production, taxonomy and antibacterial activity. J Antibiot (Tokyo) 1987; 40 (12):1677-81.

|
 IJCRR
IJCRR 





 This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License





