IJCRR - 5(2), January, 2013
Pages: 28-37
Date of Publication: 26-Jan-2013
Print Article
Download XML Download PDF
INDUCTION OF A CALCIUM DEFICIENCY IN CUTTINGS OF GRAPE VINE (Vitis vinifera L.)
Author: Bechir Ezzili
Category: General Sciences
Abstract:Rooted cuttings of grape vine, cvs. 'Black Grenache', 'Carignane' and 'Alicante Bouschet', kept at 25 \? 2\? C in the dark, I transferred it 30-days after budburst to a greenhouse at 25\?C (day), 20\?C (night) and 12 hr light. A week after, foliar growth started and normal green pigmentation appeared. 21 days later, the synthesis of anthocyanins appeared in the leaves and stems. After 28 days, calcium content, expressed as the percentage of dry matter in the shoot apex, decreased and symptoms of calcium deficiency appeared. A hypothesis was proposed to explain the mechanism inducing calcium deficiency by the pretreatment to darkness.
Keywords: Cutting one eye, pretreatment by darkness, calcium deficiency.
Full Text:
INTRODUCTION
In Tunisia, anthocyanins are mainly manufactured from grape marc. Their extraction period is limited to that of the date of harvest which lasts from 15 to 21 days according to years'. The search for other products containing these compounds can be a significant objective. We were initially interested in vine leaves (7). The results showed that certain types contain a foliar anthocyanin compounds with high levels. So their leaves could represent a promising source. In addition, and according to type of vines, the dates of harvests of the raw material, marc and leaves, are not the same ones, which allows their use. Anthocyanin extraction from the leaves would be promising. However, the search for a biotechnological process or a mechanism producing these secondary metabolites is a significant objective. We developed a technique for anthocyanin production in grape leaves (Vitis vinifera L.) cuttings (8). But, this experimental technique presents disadvantages: If we don’t harvest the leaves and the stems induced at the convenient period, yellowing and disturbances of the leaves because of a calcium disorder may occur. Calcium disorder is reviewed (18). Certain plants express this disorder: the tip-burn development in lettuce leaves (1), the bitter-pit in apples (5, 17), the blossom-end rot in pepper fruit, the apex rot potato (4) and the apex necrosis of cabbages (14), the black core of celery (19). Calcium is far from being mobile in the phloem (6, 10, 16) and its distribution depends mainly on xylem (10, 11), which can lead to disorders particularly in fabrics supplied with phloem and in fabrics which transpire little (10). According to Palzkill et al., (14, 15), root pressure can promote water and ions absorption during the night, when there is little water loss following the closing of stomata. Calcium deficiencies of the vine are induced by hydroponic culture (3). Thus, it seems that the migration of calcium is a passive phenomenon assured mainly by transpiration and to some extent by root pressure. We report here to induct a calcium deficiency from the pretreatment by total darkness.
MATERIALS AND METHODS
Grape cultivation
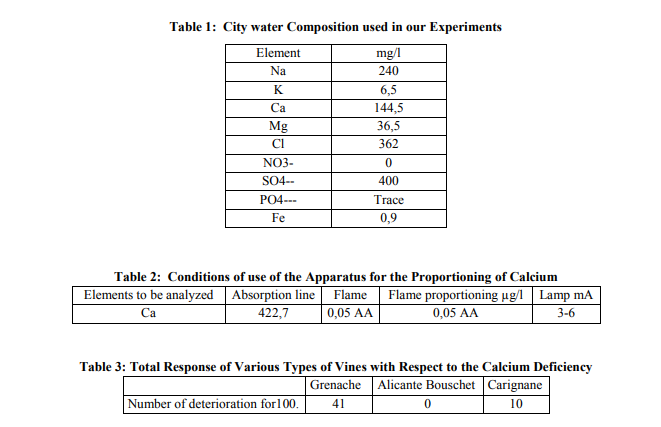
The type of vines tested are Black Grenache, Carignane and Alicante Bouschet resulting from selection massale and carried by 99 Richter, planted in 1975 in the area of El Khenguet AOC Sidi Salem delegation of Grombalia, Department of Nabeul, Tunisia, latitude 10°25’; longitude 36° 37 W; rise: 20 m. Vines were planted 2 m within and 3m between the rows, and trained to north African goblet. Fertilization, pest control and other vineyard operations were consistent with accepted commercial vineyard practices. 2000 cuttings of the cultivars (Black Grenache, Carignane and Alicante Bouschet) coming from apical fragment were cut to a length of 20 cm, disbudded so as to retain only the bud of rank 8, then planted in jars of 1 l containing city water whose composition is as presented (table 1). All cuttings were rooted with a preliminary plastering; the bud is covered with absorbent cotton, a plastic ring covers them, a band plasters is applied to the unit. The hardening of the plaster makes it possible for the bud not to strip. Routing the cutting was done then without problem. The removal of plaster then made it possible for the bud to regain its development. The cuttings were maintained at a total darkness under a relative humidity of 60% and a constant temperature of 23°C ? 2 (fig 1). By daily observations, we identified the number of buds that had reached the stage of bursting called D0. The first appearance of root occurred at the end of 35-40 days at this temperature. After appearing three roots, the buds were removed the plaster (Fig. 2). Bud bursting and branch growth in the darkness were done then without any problem (Fig.3). After 30 days, the stripped buds form then stems with highly reduced 5-6 leaves (fig. 4), characterized by the absence of chlorophyll and anthocyanin pigment. The diameter of the seedling increased. We measured it with a slide caliper. Then, we compared the section of the stem to that of the surface. If D was the diameter of the internodes n°3 in mm, the sectional area of the internodes n° 3 would be given by the formula 3.14 D2 / 4. Measures of the seedlings length and leaf area on this date of the evolution were carried out. The seedlings developed thus, exactly old 30 days, were transferred in to the greenhouse thermo regulated to 25°C day, 20°C night, 12 hours light, 12 hours darkness. After a couple of weeks, a foliar growth was noted and a normal green pigmentation appeared (Fig. 6). After 21 days, reddening of the sheets in red vines appeared (fig. 7). After 28 days in the greenhouse, calcium deficiency took place (fig. 8). They were too, of the measures of length, of the diameter of the internode’s n°3 and the leaf area are carried out. In all the cases, the length of the stem was measured. The leaf area was estimated in the following way: the leaves were detached from the seedling and were photocopied. We used the graph paper and operated with an image division, which we weighed. If P1 is the weight of the sheet image on the graph paper and P0, the weight of 100mm 2 , the surface of the sheet was then estimated by the formula 100P1 /P0 in mm 2 . For the analysis of calcium, the extraction was carried out at a stage of a 28-day stay in the greenhouse. Out of 2000 cuttings placed in the darkness, we retained 30 prototypes for each condition, gathered in a batch. The internodes of the apical and basal part were separated from the leaves which constituted 4 parts different: ISWA= Internodes subjacent with the apex, IB = Internodes of the base. SLA = subjacent leaves of the apex, LB = leaves of the base. The analyses were carried out on each part and we operated five repetitions
Proportioning of calcium
Calcium is proportioned on the product of a nitric extraction at a rate of 25 ml of nitric acid with 0,5% per 20 dry powder matter. Proportioning is done by atomic absorption in flame air-acetylene using standard photometer of flame Pye UnicamPU 9000 Philips. Certain conditions of use of the apparatus are mentioned (Table 2).
RESULTS
Total response of the seepages test screw a live treatment recommended
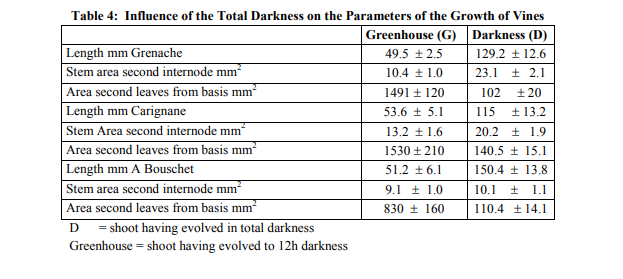
The growth of seedlings which were cultivated since the departure in the greenhouse had a normal development and did not show any red pigmentation of breaking of the branches at the final stage of their evolution. The pretreatment "darkness" involves symptoms of calcium deficiency to differing degree according to the vine type. For Black Grenache, 41% of the cuttings were affected; this effect was totally absent in Alicante Bouschet (table 3) and significantly less pronounced in Carignane (10%). The symptoms started with surface beaches which were more or less brownish and sunk into the youngest internodes. At this stage, the summit of small size could be still alive. Thereafter, the necrotic zones extended and the tips reached are desiccated. The apex died. Drying gained an apical part of the stem. We will analyse the various parameters, which are responsible for this phenomenon.
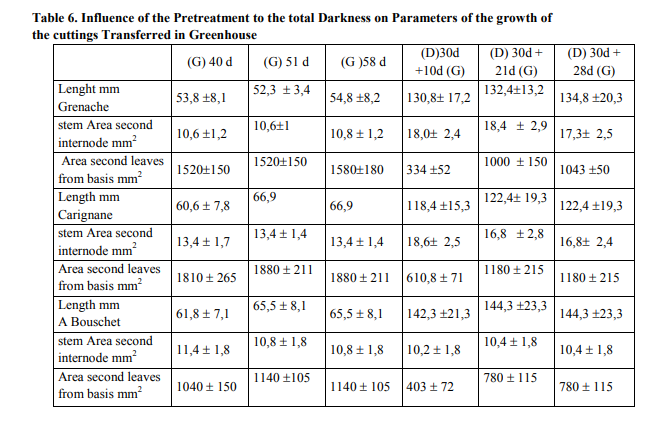
Effect of the total darkness on the characteristics of the growth of seedlings
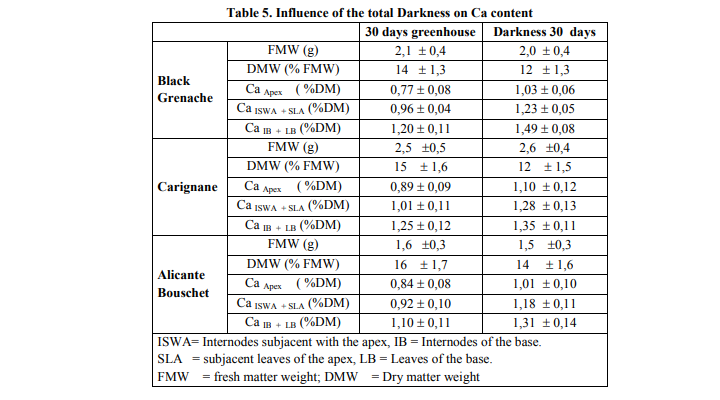
Total darkness induced an increase in the length growth of the branches of vines tested. The darkness caused as a growth in thickness which was definitely as significant as that obtained under the conditions of the greenhouse for black Grenache and Carignane, but not for Alicante Bouschet. Finally darkness reduced of 8-13 times the leaf area according to vine type, compared to the conditions of greenhouse (table 4). There were significant differences between the fresh matter weight of the seedlings cultivated under darkness and in the greenhouse for Grenache, Carignane and Alicante Bouschet. There was no difference between the fresh matter weight of the seedlings cultivated under darkness and in the greenhouse for the same type of vine. There was a significant difference between the dry matter weight of the growths cultivated in darkness and those in greenhouse for three types of vines (table 5). Darkness reduced significantly dry matter production as because of photosynthesis inhibition under this condition. There was more calcium in the basal part than the apical whatever the treatment was, in the greenhouse or in the darkness for the three types of vines. The darkness involved a reduction in the leaf area, and an increase in calcium content. These caracteristics were more pronounced in Grenache and Carignane than in Alicante Bouschet.
Effect of the pretreatment to darkness on the characteristics of the seedlings
Observation after 21 days in the greenhouse The transfer of the cuttings from darkness to the white light has not assigned to then the growth in their shoots length when comparing percentages of the increase between 30 and 51 days compared to the shoots of the cuttings cultivated directly in the greenhouse. Conversely to the length seedlings, the stem section had decreased for all groups. The etiolating shoot acquired different characters and when we returned to the greenhouse, it tended to acquire its origin characters. The transfer from the total darkness to the greenhouse for 21 days induced an increase of the leaf area of 1000%, but it did not allow recapturing the leaf area shoot average of cultivated in the greenhouse since the beginning during 51 days (Table 4 and 6). Concerning the anthocyanin compound leaves of vines, five compounds were detected: Delphinidin 3-O glucoside, Cyanidin, Petunidin, Paeonidin and Malvidin. They were presented in the free and combined form by acetic and coumaric acids (8).
Observation after 28 days in greenhouse
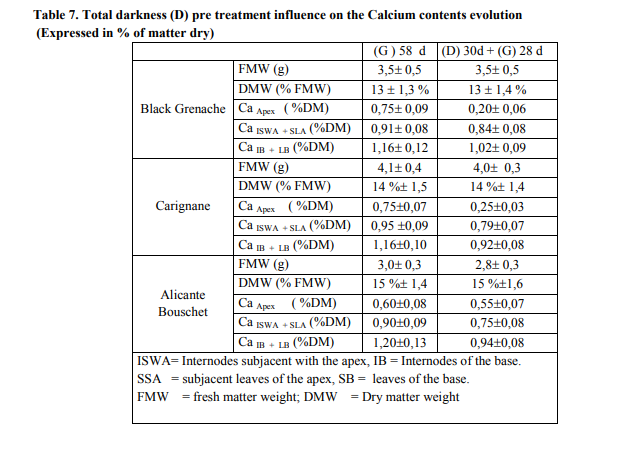
We found the same conclusions concerning the vegetative growth as those done for 21 days in the greenhouse. The principal element was the appearance of calcium deficiency symptoms,with the ultimate ones implying a cellular disintegration membrane of the apical part of the branch what disaggregates intercellular cement by dissociating the tissue structures. In the greenhouse, the growth was normal and there was no synthesis of anthocyanin compounds. There was no significant difference neither of the fresh matter weight nor the dry matter weight between the growths maintained in darkness and those in the greenhouse. There was a modification in the calcium contents particularly in the zone of the apex which decreased by 4 times for black Grenache and by 3 times for Carignane. For Alicante Bouschet, the calcium content was not significantly affected (table7). In the darkness, the dry matter weight is 12% for Grenache and Carignane and 14 % for Alicante Bouschet (table 5). When the growths condition was changed to the greenhouse, the dry matter weight became 13% for Grenache, 14% for Carignane and 15 % for Alicante Bouschet (table 7). There was not variation in the change of the matter fresh weight between the growths exposed directly in the greenhouse and those pretreated by the total darkness. The transfer of the batch from total darkness to the greenhouse had not affected the calcium contents of the basal part of the growth (table 7). In our program two periods were followed: the first was characterized by total darkness during 30 days (phase I) and the second by the transfer in to the greenhouse during 28 days (phase II). In phase I, the stomata closing and the reduced surface of the leaves suggested that water loss by transpiration was weak and the root pressure was dominant. In phase II was characterized by the transfer of the batch from total darkness to the greenhouse and mainly by a resumption of the leaf growth, the transpiration would over ridded it this time on the root pressure and the calcium deficiency was observed. In addition, under temperature conditions, moisture and stationary photoperiod, different calcium absorption happens in the vines stems according to vine type. For black Grenache the calcium contents expressed as the percentage of the dry matter are a quarter, for a one third for Carignane when there was appearance of the necrosis apical symptoms. On the other hand, differences in calcium contents are not significant between treated and witnesses; whereas growth in width stems was practically the same in treated as in the greenhouse, although the foliar growth in the treated seedlings was reduced in the same way as that of two other vine types.
DISCUSSION AND CONCLUSION
The calcium transport for the plants was studied (9, 12, 13, 20). We retain the following data: - Water penetrates through the hair absorbants, gets to the vessels by the bark and the central cylinder: It follows the way of the apoplasm, the symplasm and transcellular transport. - The migration of calcium at long distance is assured passive fact a mainly by transpirant leaves. The root pressure is a way additional or a replacement of transpirant leaves. It determines an ascending transport. - The crude sap distribuates calcium to the various parts of the plant. The calcium content in the elaborate sap is weak. Calcium is not mobile, it is not distributed the oldest leaves towards young. - The apex is supplied with calcium by migration on sites of exchanges. Mitosis and the growth of cells generate new sites of fixation at the level of the plasmalemn and average plate that calcium will occupy by cationic reaction of exchange while following the apoplasmic way. On the basis of these data, we propose an explanation of the manifestation of the calcium deficiency. The pretreatment of the seedlings in total darkness induced etiolation responsible for three vine types, a lengthening of the stem, an abnormal thickness growth and a considerable reduction of the leaf area as well as a blocking of the anthocyanins synthesis. These conditions are due to phototropism and far UV (2). The ascending transport of calcium is done mainly by the root pressure. This means of transport is thus able to feed the growth equipped with their leaves for their calcium apex. It is then the means of transport at long distance from this element under this condition. However it is generally allotted a palliative or complementary function to transpiration (12, 13). In the greenhouse, the growth is normal and there is no synthesis of anthocyanin compounds, the transport of calcium is done by transpiration for the diurnal period and probably by the root pressure at the night. For the vine, the root pressure is effective only if the transpiration feeds it. During the night, It contributes to the continuation of the routing of the calcium imported beforehand in the branches by the transpirant leaves for the diurnal period. Radicular flow would be a temporary vehicle relay of calcium requiring a regular restocking. The passage from total darkness to the greenhouse is followed by a resumption of the seedlings (leaf biomass), a reduction of the stem section and an induction of the biosynthesis of anthocyanins. The physiological state is regained by the conditions of the greenhouse. When the seedlings undergo a preprocessing the darkness followed by that of the greenhouse, the root pressure initially generates an ascending transport of calcium and ensures calcic food for the whole seedlings. During the first days, after the transfer into the greenhouse, the apex is probably supplied with calcium by the root pressure. As the leaf area of the base seedlings increases, the top of the growth is insufficiently fed. The flow of calcium is directed preferentially towards the base, and the symptoms of deficiency appear at the top. As soon as the transpiration of the basal sheets becomes intense, there are deficiency on the apex and appearance of the symptoms of calcium disorder. The migration of water leaves of the base have a relatively large leaf area, the transpiration takes over. The diversion of the flow of calcium towards the mature leaves is probably at the origin of the symptoms of calcium deficiency, which appear on the level of the apical part of the cutting.
References:
1. Barta, D. J., Tibbits, T. W. (2000). Calcium localization and Tipp burn development in lettuce leaves during early enlargement. J. Am. Soc. Hort. Sci. 125: 294-298.
2. Briggs, W.R., Mosinger, E., Schafer, E. (1988). Phytochrom regulation of greening in barley- Effets on chlorophyll accumulation. Plant Physiol. 86: 435-440.
3. Bruzau, F., Juste, C. et Poujet, R. (1968). Mise en évidence d'une carence en calcium chez la vigne cultivée en solution nutritive. C.R.A.S.S. PARIS. 266 : 116-118.
4. Dekock, P.C, Dyson, P.W, Hall, A. et Grabowska, F. (1975). Metabolic changes associated with calcium deficiency in potato sprouts. Potato Research. 18: 573-581.
5. Drazeta, L. A., Hall A. J. Volz, R., Jameson, P.E. (2004). Causes and effect of change in xylem functionality in apple fruit. Ann. Bot. 93: 275-282.
6. Epstein, E. (1973). Flow in the phloem and the immobility of calcium and boron : a new hypothesis in support of an old one. Experimentia. 29: 133-134.
7. Ezzili, B., Habib, J., Darne, G., et Chemli, R. (1997). lnfluence de la date de prélèvement sur les teneurs en anthocyanes et en éléments minéraux du cépage Alicante Bouschet cultivé à El khenguet (Tunisie); Bull. OIV. 70: 795-796.
8. Ezzili, B; Darne, G. et Bejaoui, M. (1999). Mise au point d’une technique de production d’anthocyanes dans les feuilles de boutures à un œil de Carignan (Vitis vinifera L.) cultivées au laboratoire. J. Int. Sci. Vigne. Vin. 33: 9-18.
9. Ferguson, I.B. et Bollard, E. G. (1976). The movement of calcium in woody stems. Ann. Bot. 40: 1057-1065.
10. Hocking, P.G., Pate, J.S., .Atkins, A. et Sharkey, P.J. (1978). Diurnal patterns of transport and accumulation of minerals in fruiting plants of Lupinus augustifolius. Ann. Bot. 42 : 1277-1290.
11. Glad, O., Regnard, J.L., Queron, Y., Brun, O., Morot-Gaudry, J.F. (1992). Flux and chemical composition of xylem exsudates from Chardonnay grapevine: Temporal evolution and effect of recut. Am. J. Enol. Vitic. 43: 275-282.
12. Kirkby, E.A., Pilbeam, D.J. (1984). Calcium as nutrient. Plant cell and environment, 7: 397-405.
13. Marschner, H. (1995). Mineral Nutrition of Higher Plants. Academic Press, Amsterdam, 2 ed edition
14. Palzkill, D.A., Tibbits, T.W., Williams, P.H. (1976). Enhancement of calcium transport to inner leaves of Cabbage for prevention of Tipburn. J. Amer. Soc. Hort. Sci., 101: 605- 648.
15. Palzkill, D. A., Tibbits T. W., (1977). Evidence that root pressure is required for calcium transport to head leaves of Cabbage. Plant Physiol.
16: 682-694. 16. Raven, A. (1977). H+ and Ca++ in phloem and symplast : Relation of relative immobility of the ions to the cytoplasmic nature of transport paths. New Phytol. 79: 465-480.
17. Saure, M.C. (1996). Reassessment of the role of calcium in development of bitter pit in apple .Aust. J. Plant. Physiol. 23: 237-243.
18. Saure, M.C. (2005). Calcium translocation to fleshy fruit: its mechanism and endogenous control. Scientia Horticulturae 105: 65-89.
19. Shear, C. B. (1975). Calcium related disorders of fruits and vegetables. Hort Science, 10:361-365.
20. Van De Geijn S. C. Smeulders E. (1981). Diurnal changes in the flux of calcium toward meristems and transpiring leaves in Tomato and Maize plants. Planta, 151: 265- 271.

Legende of figures
Procedure for the induction of a calcic deficiency at the laboratory
1 Bud is plastered in order to induce roots.
2 When there are at least 3 roots the bud is removed the plaster.
3 bud bursting in darkness
4 Stage six very reduced sheets stage 30 days in darkness
5 Stage 30 days in darkness and in greenhouse. On the left, shoots in total darkness can have 7, 8, 9, 12 very reduced a pale yellow sheets. On the right, cuttings are cultivated in greenhouse give normal axes with spread out and rather large sheets. To note here the difference of lengthening of the branches and growth of the appreciable sheets.
6 Cutting transferred from the darkness to the greenhouse during 15 days. Note here the beginning of colouring of the stems.
7 Cutting transferred from the darkness to the greenhouse during 22 days. To note Curve of the stem of approximately 45°C and sunk red colouring of the carrying branch.
8 Cutting transferred from the darkness to the greenhouse during 30 days. To note the curve of an angle equal to 135°C and beginning of the drying of the apex which starts approximately at a point located at the third of the branch.
|
 IJCRR
IJCRR 





 This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License





